
13.2: Fates of Pyruvate under Anaerobic Conditions- Fermentation
- Page ID
- 15006


\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)-
Explain the Purpose of Fermentation:
- Define fermentation as an anaerobic process that enables ATP production when oxygen is limited, emphasizing the critical role of glycolysis and the recycling of NADH to NAD⁺.
-
Describe NAD⁺/NADH Recycling in Fermentation:
- Explain why recycling NADH to NAD⁺ is essential for sustaining glycolysis under anaerobic conditions and compare how this recycling occurs via aerobic pathways (e.g., the citric acid cycle) versus anaerobic fermentation.
-
Differentiate Between Lactate and Ethanol Fermentation:
- Contrast the two major anaerobic fermentation routes: lactate fermentation (conversion of pyruvate to lactate by lactate dehydrogenase) and ethanol fermentation (pyruvate decarboxylation to acetaldehyde followed by reduction to ethanol by alcohol dehydrogenase).
-
Understand Tissue-Specific Fermentative Pathways:
- Identify the physiological contexts in which each fermentation type occurs—for example, lactate production in mammalian muscle and red blood cells, and ethanol production in yeast and certain fish species.
-
Explain the Cori Cycle and Its Importance:
- Describe the Cori cycle, including how lactate produced in muscle is transported to the liver for conversion back to pyruvate and subsequently to glucose via gluconeogenesis, highlighting its role in maintaining energy homeostasis.
-
Analyze Enzyme Structure and Regulation in Fermentation:
- Detail the structural and mechanistic aspects of key enzymes (e.g., lactate dehydrogenase, pyruvate decarboxylase, alcohol dehydrogenase), including their isozyme variations, active site configuration, and allosteric regulation.
-
Discuss Ethanol Metabolism in Humans:
- Outline the sequential metabolism of ethanol in the liver by cytosolic alcohol dehydrogenase (ADH) and mitochondrial acetaldehyde dehydrogenase (ALDH2), and explain how disruptions in these pathways (e.g., accumulation of acetaldehyde) contribute to toxicity.
-
Evaluate the Effects of Alcohol Consumption:
- Assess the concept of hormesis in alcohol consumption, discussing how moderate drinking may be cardioprotective while excessive intake leads to detrimental outcomes such as liver damage, altered gene expression, and increased cancer risk.
-
Link Fermentative Metabolism to Systemic and Cellular Health:
- Connect changes in NADH/NAD⁺ ratios during ethanol metabolism to broader metabolic effects, including downregulation of glycolysis and the Krebs cycle, lactic acidosis, and hypoglycemia.
-
Examine Therapeutic Strategies for Alcohol Abuse:
- Understand the mechanism of disulfiram as an inhibitor of ALDH, the rationale behind its use in treating alcoholism, and its role in promoting abstinence through the accumulation of toxic acetaldehyde.
These learning goals are intended to provide a comprehensive framework for understanding fermentation’s biochemical basis, its integration with cellular metabolism, and its physiological and clinical implications.
Introduction to Fermentation
Fermentation occurs when a fuel molecule is broken down anaerobically (or without oxygen) to meet ATP demands. One of the most notable pathways to utilize fermentation is glycolysis, which we have just described. During glycolysis, glucose is converted to pyruvate, yielding 2 ATP energy-rich molecules. This occurs in anoxic conditions (or without the requirement of oxygen). However, the process of glycolysis cannot be sustained if the end product is pyruvate. This is because the production of pyruvate also yields two molecules of reduced NADH in addition to the production of the two ATP molecules. The pool of available NAD+/NADH is limited within the body, and thus, this electron acceptor/donor molecule must continually be recycled for metabolic pathways to remain functional. For example, if NADH is not oxidized back into NAD+ in a timely manner, the glycolytic pathway can slow. Therefore, to be able to maintain the process of energy generation through glycolysis, the pool of NADH needs to be converted from the reduced form back into the oxidized form (NAD+), where it can then accept electrons from intermediates in the glycolytic pathway. This process of recycling NADH to NAD+ can occur aerobically through the Citric Acid Cycle, which is described in more detail in Chapter 16, or it can be processed via anaerobic fermentation (Figure \(\PageIndex{1}\)).
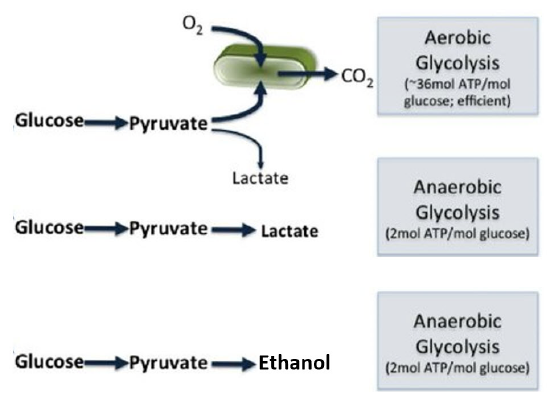
Figure \(\PageIndex{1}\): Fates of Pyruvate via Aerobic or Anaerobic Pathways.
In the 1860s, Louis Pasteur first described fermentation very narrowly as the process yeast uses to convert glucose into ethanol when grown in the absence of air. Subsequently, it was discovered that other microorganisms could convert pyruvate to lactate (or lactic acid) instead of ethanol during anaerobic respiration. These microorganisms became known as lactic acid bacteria and are currently utilized heavily within the food industry to produce a wide array of fermented food products, including cheese, yogurt, and sauerkraut, to name a few. As the enzymes within the glycolytic pathway were discovered, it became apparent that muscle tissue could also engage in anaerobic respiration, producing lactate. Thus, while fermentative processes are most often described in microbial organisms, the definition of fermentation has since been broadened to include any enzymatic, energy-yielding pathways that occur in the absence of oxygen, including the production of lactate in muscle tissue of animals during the glycolytic process.
 Industry is turning to microbes to produce new food products such as animal-free proteins and novel foods, drugs, household products, and chemical feedstocks to produce commercial products like biofuels and even plexiglass, to name just a few. Check out Chapter 32.18: Turning Trees into Plexiglass: Synthetic Biology For Production of Green Foods and Products for more details!
Industry is turning to microbes to produce new food products such as animal-free proteins and novel foods, drugs, household products, and chemical feedstocks to produce commercial products like biofuels and even plexiglass, to name just a few. Check out Chapter 32.18: Turning Trees into Plexiglass: Synthetic Biology For Production of Green Foods and Products for more details!
Synthetic biology, using existing and modified metabolic pathways in microorganisms to accomplish these tasks, is revolutionizing manufacturing to make it greener, more sustainable, and with a lower carbon footprint. One major goal is to reduce CO2 emissions to reduce global warming and its consequences. When the pathways are highly engineered and tailored to specific goals, it is often called precision fermentation. These microbial pathways can also be anaerobic or aerobic, so the original definition of fermentation involving anaerobic reactions has been expanded.
NADH recycling to NAD+, typically in anaerobic fermentation reactions, occurs by two different routes:
- lactate fermentation (Figure \(\PageIndex{1}\) in bacteria and fungi, such as Lactobacillus, Leuconostoc mesenteroides, Pediococcus cerevisiae, Streptococcus lactis, and Bifidobacterium bifidus) and, of course, the muscles of animals. Yogurt is the product of the anaerobic fermentation of milk by microorganisms such as Lactobacillus delbrueckii subsp. bulgaricus and Streptococcus thermophilus.
- ethanol fermentation in yeast and some bacteria, such as Zymomonas mobilis.
We will focus on the details of these two systems here.
Lactate Fermentation
In lactate fermentation, pyruvate is converted to lactate by the enzyme lactate dehydrogenase. In the process, NAD+ is regenerated. The reaction catalyzed by lactate dehydrogenase (LDH) is shown in Figure \(\PageIndex{2}\).

Figure \(\PageIndex{2}\): Reaction catalyzed by lactate dehydrogenase
It is named for the reverse reaction, which uses NAD+ as an oxidizing agent, as with other dehydrogenases. The reaction is reversible with the ΔG0 for conversion of pyruvate to lactate of -3.76 kcal/mol (-15.7 kJ/mol). Under anaerobic conditions, when glycolysis is the major source of ATP, pyruvate levels increase, further driving the reaction towards lactate formation and NAD+ regeneration so glycolysis can continue.
The enzyme is found in the cytoplasm, but a mitochondrial form also exists. It is most abundant in muscle, liver, and kidney, and it is also found in erythrocytes. Interestingly, mature red blood cells are enucleated and do not contain any mitochondria. Their lifespan is limited to approximately two weeks. During this time, their primary energy resources are generated through anaerobic fermentation via the glycolysis-lactate pathway.
The active enzyme is a tetramer of various compositions of two different subunits, the heart (H) and muscle (M) forms. The quaternary structures contain five different isozyme forms containing the H and M subunits. LDH-1, found most abundantly in the heart, is a tetramer of 4H subunits (H4). The other forms are as follows: LDH-2 (H3M, prevalent in red blood cells), LDL-3 (H2M2, prevalent in lungs), LDH-4 (HM3, prevalent in the kidney), and LDH-5 (M4, prevalent in muscle).
During anaerobic metabolism, lactate is produced by muscle tissue and released into the bloodstream, where it can travel back to the liver. Once in the liver, lactate is converted back into pyruvate and can be utilized to produce glucose through a pathway called gluconeogenesis. The liver can export glucose into the blood. There, it is taken up by the muscle and used for ATP production. This cycle is called the Cori cycle, illustrated in Figure \(\PageIndex{3}\).
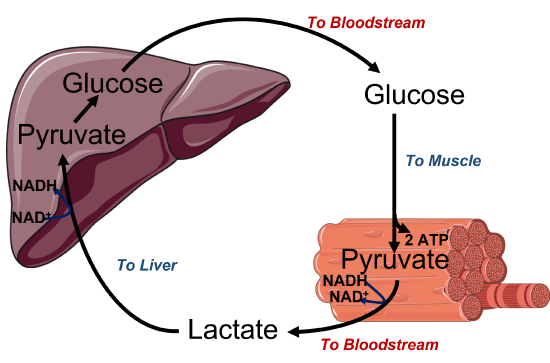

Figure \(\PageIndex{3}\): Cori cycle. The Upper diagram is a cartoon image of the Cori cycle, and the lower diagram demonstrates the recycling of NAD+/NADH in different locations within the body. Figure modified from Servier Medical Art
Notably, animal anaerobic metabolism can only be sustained for short periods due to its high energy demand. Aerobic respiration is required to maintain an adequate ATP supply. However, the production of lactate by certain tissues, such as white or fast-twitch muscle cells, occurs regularly and releases lactate into the bloodstream, which can be taken up and used as an energy source by neighboring tissues, such as red, slow-twitch muscle. Brain tissue can also effectively use lactate as an energy source. Within these 'consumer' tissues, lactate is converted back into pyruvate using the mitochondrial LDH enzyme. It can be converted to pyruvate and used in aerobic respiration to produce high levels of ATP.
Thus, lactate production can be considered a response during metabolic stress, such as intense cardiovascular exercise. It has also been noted that after an injury or head trauma, the activation of epinephrine will cause an increase in lactate production, and blood levels of lactate will increase. Thus, it has been hypothesized that lactate may play a role in the repair of damaged tissue. Clinical trials and experiments are currently underway to determine if lactate can help in the healing and recovery process for conditions such as traumatic brain injury, myocardial infarction, and sepsis. Early studies have shown that lactate can increase the production of Brain-Derived Neurotropic Factor (BDNF), which supports neuronal growth, providing further support for the role of lactate in recovery and repair.
Spectroscopy analyses show that the geometry and electrostatics of the active sites of LDHs from different organisms are essentially the same. The mechanism for the reverse oxidation of lactate by NAD+ by spiny dogfish lactate dehydrogenase (LDH) is shown in Figure \(\PageIndex{4}\)

Figure \(\PageIndex{4}\): Mechanism for the reduction of lactate by NAD+ by spiny dogfish lactate dehydrogenase (LDH)
Lactate is shown with a deuterium (D), which moves as a deuteride to NAD+ to form NADH, to illustrate the stereochemistry of the reaction. In the reverse reaction, reducing pyruvate to lactate (to regenerate more NAD+ so anaerobic glycolysis can continue), the deuterium (or the proR H of non-deuterated NADH) is removed.
Figure \(\PageIndex{5}\) shows an interactive iCn3D model of dogfish M4 apo-lactate dehydrogenase (1LDM).
.png?revision=1&size=bestfit&width=680&height=371)
 Figure \(\PageIndex{5}\): Dogfish M4 apo-lactate dehydrogenase (1LDM).(Copyright; author via source). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...FBzuqJ2gEPUHM8
Figure \(\PageIndex{5}\): Dogfish M4 apo-lactate dehydrogenase (1LDM).(Copyright; author via source). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...FBzuqJ2gEPUHM8
Only the monomer of the active tetramer is shown. Oxamic acid (labeled) is structurally similar to lactate/pyruvate and is a competitive inhibitor of the enzyme. His193, the active site general base/acid, is near Arg 169, and also to Asp 166 and Asn 138, which are hydrogen bonded to the inhibitors.
LDH exists in two major conformational states, the T (inactive) and R (active) states, as seen with other allosteric proteins. Figure \(\PageIndex{6}\) shows an interactive iCn3D model of the T state (2ZQY) and R state (2ZQZ) of Lacticaseibacillus casei L-lactate dehydrogenase.
.png?revision=1&size=bestfit&width=415&height=380)
Figure \(\PageIndex{6}\): T state (2ZQY) and R state (2ZQZ) of Lacticaseibacillus casei L-lactate dehydrogenase. (Copyright; author via source). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...zjrtVYjLXdhDp8
Toggle between the magenta T and cyan R states using the "a" key. Orient the proteins to see the best view of the allosteric changes caused by binding regulators. The spheres show Arg 171, whose disposition changes significantly in the T and R states. Pyruvate (a reactant/product) activates the enzyme (conversion of T to R state) and is considered a homotropic regulator since it is a substrate. This is similar to hemoglobin's allosteric "activation" by its ligand O2, which preferentially binds to the R state and promotes the T-to-R state transition. Fructose 1,6-bisphosphate, a glycolytic intermediate, also activates the Lacticaseibacillus casei L-lactate dehydrogenase, and since it is not a reactant/product of the enzyme, it is considered a heterotropic allosteric regulator. Hence, the substrate concentration (pyruvate) and NADH levels effectively regulate the enzyme. High concentrations of ethanol consumption lead to high levels of NADH through the activity of alcohol dehydrogenase. High levels of NADH would lead to increased lactate production as well.
Ethanol Fermentation in Yeast
In this process, pyruvate is decarboxylated first to acetaldehyde by the thiamine pyrophosphate (TPP)-requiring enzyme pyruvate decarboxylase. The resulting product, acetaldehyde, is then reduced by NADH to form ethanol by the enzyme ethanol dehydrogenase in a process that reforms NAD+. Yeast is a facultative (not obligate) anaerobe in that it can produce energy by glycolysis and ethanol fermentation in the absence of oxygen. Figure \(\PageIndex{7}\):. Of course, in the presence of oxygen, the pyruvate produced from glycolysis in yeast is preferentially converted to acetyl-CoA, which enters the citric acid cycle and oxidative phosphorylation pathways to maximize ATP production.
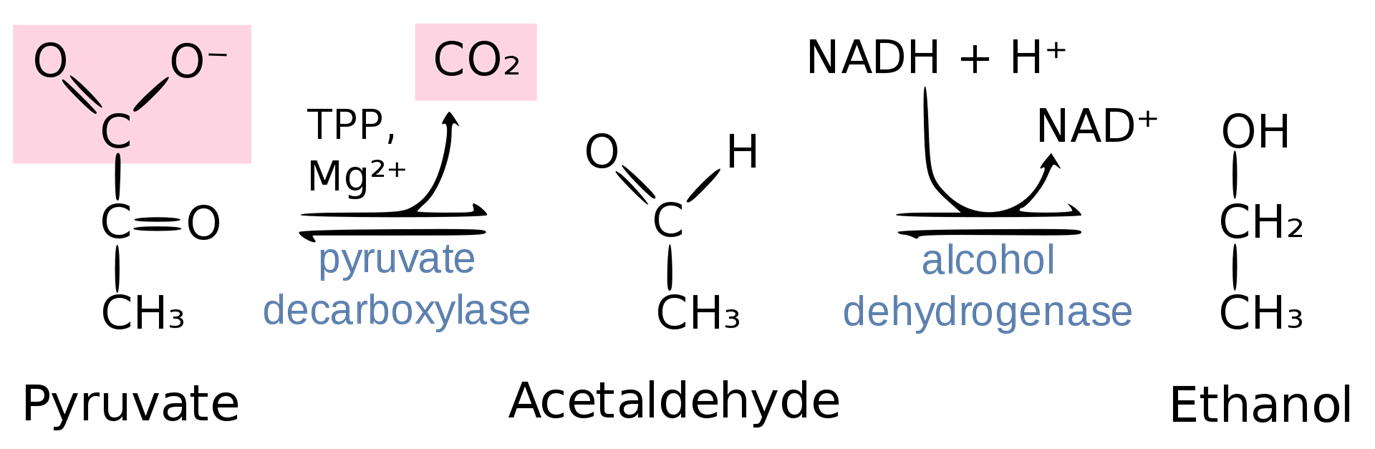
Figure \(\PageIndex{7}\): Summary of Ethanol Fermentation in Yeast. Figure from Thomas Baldwin
Pyruvate decarboxylase
Pyruvate decarboxylase catalyzes the first step in the alcoholic fermentation pathway, converting pyruvate into acetaldehyde and carbon dioxide. Interestingly, in addition to yeast, some fish species, such as goldfish and carp, have a homologous enzyme that allows for ethanol production when oxygen is scarce. Pyruvate decarboxylase is tetrameric, occurring as a dimer of dimers with two active sites shared between the monomer subunits of each dimer. The decarboxylation reaction requires two cofactors, thiamine pyrophosphate (TPP) and magnesium (Figure \(\PageIndex{8}\)). Within the active site, the acidic Glu-477 and Glu-51 residues and the Mg2+ cofactor interact with and stabilize the TPP cofactor. The aminopyrimidine ring on TPP acts as a base and enables the formation of the TPP nucleophile by removing the C2 proton. The protonation of Glu-51 stabilizes this reaction. The nucleophilic attack of pyruvate causes the release of carbon dioxide. The acetaldehyde intermediate is still covalently attached to the TPP cofactor following the release of carbon dioxide.

Figure \(\PageIndex{8}\): Part 1 Pyruvate Decarboxylase mechanism - decarboxylation
Once carbon dioxide diffuses away from the active site, the double bond of the enol intermediate abstracts a proton from Asp-28, which is stabilized by a neighboring His-115 residue. TPP can then serve as a good leaving group during the formation of the carbonyl functional group and causes the release of acetaldehyde (Figure \(\PageIndex{9}\):).

Figure \(\PageIndex{9}\): Part 2 Pyruvate Decarboxylase mechanism - acetaldehyde generation
Figure \(\PageIndex{10}\) shows an interactive iCn3D model of the thiamin diphosphate-dependent enzyme pyruvate decarboxylase from the yeast Saccharomyces cerevisiae (1PVD).
.png?revision=1&size=bestfit&width=583&height=376)
Figure \(\PageIndex{10}\): Thiamin diphosphate-dependent enzyme pyruvate decarboxylase from the yeast Saccharomyces cerevisiae (1PVD).(Copyright; author via source). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...DMM4gjodX4q1G9 TTP is shown in spacefill. The enzyme is a homotetramer.
Alcohol dehydrogenase (ADH)
Alcohol dehydrogenase enzymes catalyze the interconversion of aldehydes or ketones with alcohol functional groups. In humans and other animals, they are utilized to break down ingested alcohols and participate in the biosynthesis of numerous metabolites. There are at least five classes of alcohol dehydrogenases that use NAD+ as an oxidizing agent to convert alcohols to aldehydes or ketones. We are mainly concerned with the yeast ADH, a member of Family I, that is involved in ethanol fermentation. Figure \(\PageIndex{11}\) shows the reaction catalyzed by alcohol dehydrogenase.

Figure \(\PageIndex{11}\): Reaction catalyzed by alcohol dehydrogenase
Figure \(\PageIndex{12A}\) below shows a mechanism for yeast alcohol dehydrogenase (ADH1). ADH1 requires zinc as a metal cofactor and utilizes NADH as the reducing agent in converting acetaldehyde into ethanol.

Figure \(\PageIndex{12A}\): Simplified catalytic mechanism of ADH reactions. Upon binding of both substrates (e.g., NAD(P)+ and alcohol; 0->1->2), a hydride transfer occurs from the alcohol-carbon atom to the oxidized nicotinamide moiety, yielding the Zn-coordinated carbonyl product and NAD(P)H (3). Both can dissociate from the active site, yielding apo-ADH (0). Alternatively, only NAD(P)H stays bound, and the reduced ADH can undergo a reductive conversion (1’->2’-> 3’-> 1). Amanda Silva de Miranda, et al. Front. Catal., 10 May 2022. Sec. Biocatalysis. https://doi.org/10.3389/fctls.2022.900554. Creative Commons Attribution License (CC BY).
Figure \(\PageIndex{12B}\) shows key amino acids in the active site of yeast ADH1. Note that in this figure, two cysteines are the coordinating ligands.

Figure \(\PageIndex{12B}\): Abbreviated mechanism for yeast alcohol dehydrogenase (ADH1)
If you look at the reverse reduction reaction, the acetaldehyde (or a ketone for 20 alcohols) carbonyl is sp2 hybridized and planar, so the hydride could be added to either of the two faces, re or si, of the plane (using the same rules used to define R or S enantiomers). Likewise, the methylene carbon atom with two hydrogens, one of which is transferred as a hydride, is prochiral. Hence, four different stereochemical hydride transfer pathways are possible, as shown below in Figure \(\PageIndex{13A}\). Hydride transfers from the prochiral aldehyde/ketone si-face result in (R)-configured alcohols, whereas hydride attacks from the re-face yield (S)-alcohols. In both cases, the hydride transferred can stem from either the re- or si-face of the nicotinamide ring.

Figure \(\PageIndex{13A}\): Possible stereochemical courses of the hydride transfer from NAD(P)H to the ketone. Attacks from the si-face of the ketone result in (R)-alcohols (E1 and E2), whereas hydride attacks from the re-face (E3 and E4) result in (S)-alcohols.
ADHs catalyzing hydride addition from the re-face of the ketone (or abstraction of a hydride from (S)-alcohols) are termed Prelog-selective ADHs, whereas those ADHs attacking from the si-face (or abstracting a hydride from the (R)-alcohol) are termed anti-Prelog ADHs. Different ADHs are available for the different reaction stereochemistries.
The stereochemistry for the reaction of ADH1 with ethanol is shown in Figure \(\PageIndex{13B}\) below. Deuterium (D) is shown in the figure to illustrate the reaction's stereochemistry better. With the deuterium label, the carbon is now chiral, and the enantiomer shown is the R isomer. The enzyme removes the corresponding H in the undeuterated and prochiral form of ethanol, as shown in Figure \(\PageIndex{13B}\) below.

Figure \(\PageIndex{13B}\): Stereochemistry of human alcohol dehydrogenase reaction.
Figure \(\PageIndex{14}\) shows an interactive iCn3D model of the Yeast alcohol dehydrogenase (ADH I) with bound substrate analogs- ADH1 (4W6Z).
.png?revision=1&size=bestfit&width=465&height=382)
Figure \(\PageIndex{14}\): Yeast alcohol dehydrogenase structure with bound substrate analogs (4W6Z). (Copyright; author via source). Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...feE7toF6wU6MB9
All four monomers of the homotetramer are shown. The light gray subunit has the key catalytic and binding residues shown in sticks and labeled. The numbering of key residues differs slightly from the mechanism shown above. The cofactor analog, nicotinamide-8-iodo-adenine-dinucleotide, is shown in sticks, and the substrate analog, trifluoroethanol, is shown in spacefill.
Only two subunits (a homodimer) have both bound cofactor and substrate. The substrate trifluoroethanol in these subunits is ligated to the catalytic Zn2+ through its oxygen, with the other ligands provided by the side chains of Cys 43, Cys 153, and His 66. The other two monomers (the other homodimer) have a different conformation and use Glu 67 to coordinate the Zn2+ instead of His 66, and no substrate is bound. This may be an intermediate in the process that displaces water bound to Zn2+ with the substrate.
Alcohol Metabolism in Humans:
When ethanol is consumed, it is oxidatively metabolized primarily in the liver. Cytosolic Alcohol Dehydrogenase (ADH) and mitochondrial Acetaldehyde Dehydrogenase 2 (ALDH2) are the main enzymes involved in this metabolic pathway, first converting ethanol to acetaldehyde and then acetaldehyde to acetate (Figure \(\PageIndex{15}\):). Liver mitochondria have a limited capacity to use the acetate in the Kreb cycle because the enzyme needed to convert acetate to acetyl-CoA (acetyl-CoA synthase 2) is almost absent in the liver but is abundant in the heart and skeletal muscles. Thus, most of the acetate resulting from ethanol metabolism escapes the liver into the blood circulation and is eventually metabolized to CO2 by way of the Krebs cycle in cells with mitochondria that contain enzymes to convert acetate to acetyl-CoA, such as the heart, skeletal muscle, and brain.
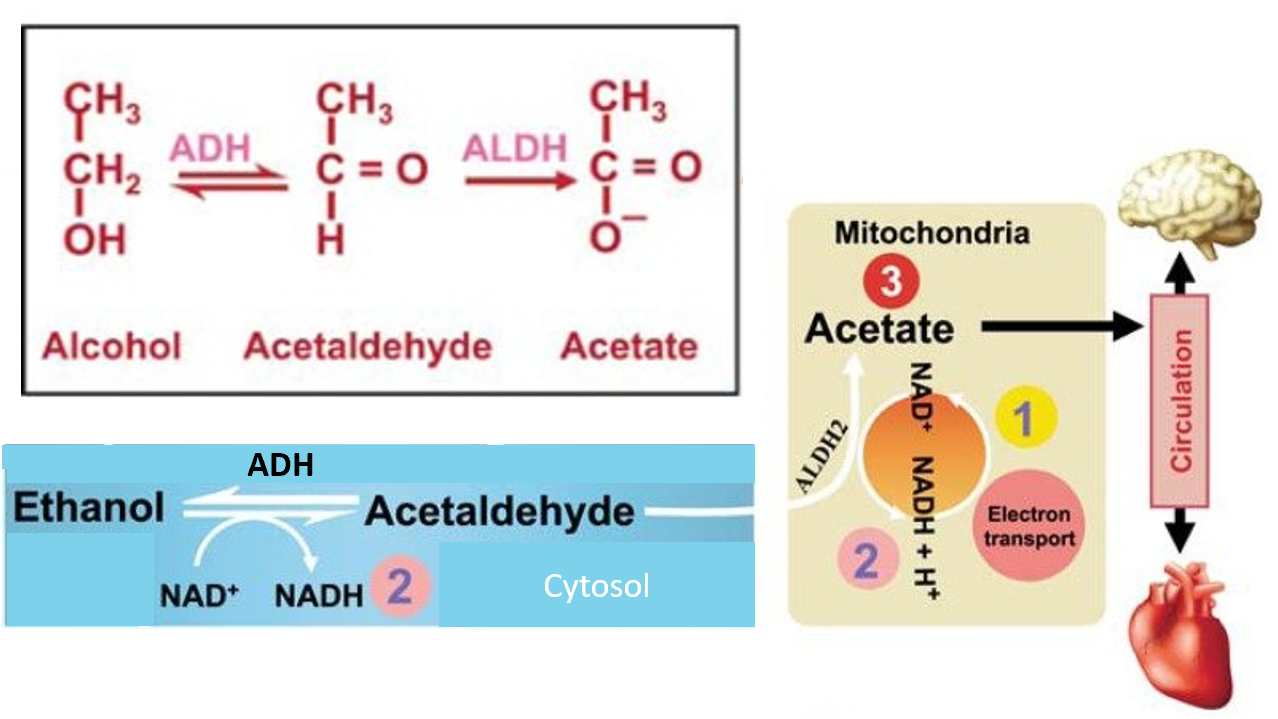
Figure \(\PageIndex{15}\): Summary of Ethanol Metabolism in Humans. Figure modified from Zakhari, S. (2013) Alcohol Research: Current Reviews 35(1):6-16
Like many substances, alcohol can be both beneficial and detrimental, depending on the quantity and frequency of consumption. This is known as a hormetic response (Figure \(\PageIndex{16}\):). At low doses (up to two daily drinks for men and one daily drink for women), the consumption of alcohol may be cardioprotective. At higher consumption rates, the detrimental effects of alcohol consumption become apparent and include cardiotoxicity, liver damage, and increased cancer risk, not to mention the debilitation that can accompany addiction.

Figure \(\PageIndex{16}\): Alcohol Consumption and Hormesis. Figure from Adamson, S.S., et al (2017) Translational Medicine of Aging 1:18-23.
The graph in Figure \(\PageIndex{17}\) exemplifies the hormetic nature of alcohol consumption. The data show that having one drink per day lowers overall mortality, but after that, there is a sharp rise.

Figure \(\PageIndex{17}\): Risk of Mortality and Alcohol Consumption. Tian, Y., Liu, J., Zhao, Y. et al. Alcohol consumption and all-cause and cause-specific mortality among US adults: prospective cohort study. BMC Med 21, 208 (2023). https://doi.org/10.1186/s12916-023-02907-6. Creative Commons Attribution 4.0 International License. http://creativecommons.org/licenses/by/4.0/.
Dose–response relationship between alcohol consumption and risk of mortality from all causes. A nonlinear relationship of current alcohol consumption (as a continuous variable) with all-cause mortality (p < 0.05 for the nonlinear test), using maximally adjusted estimates (adjusted for sex, age, race/ethnicity, education, marital status, body mass index, physical activity, smoking, and physician-diagnosed diseases (heart disease, stroke, cancer, diabetes, hypertension, asthma, emphysema, and chronic bronchitis). That indicates that alcohol consumption had a J-shaped association with risk of all-cause mortality. HR, hazard ratio.
The Centers for Disease Control estimates that alcohol abuse leads to approximately 80,000 deaths annually in the United States and that up to 40% of deaths related to liver disease are caused by alcohol abuse. Harvard Medical School has evaluated thousands of studies and developed a method for alcohol risk assessment. They categorize low-risk drinking behavior for men as no more than four drinks in a single day or 14 drinks in a week, and no more than three drinks in a single day or a total of seven drinks in a single week for women. Women have lower drinking tolerance due to their smaller size but also due to metabolic differences. Women have less ADH and typically higher fat levels, which cause ethanol to disperse and be retained longer. Addiction and damage to the liver tend to occur more quickly in women as well. Harvard ranks people at increased risk if their drinking is above either the single-day or the weekly limit (which is estimated at 29% of drinkers). High-risk drinkers break both the daily and weekly limits (estimated at 14% of drinkers).
During ethanol metabolism, when circulating ethanol is in the millimolar range, acetaldehyde is in the micromolar range, and acetate is in the millimolar range (Figure \(\PageIndex{18}\):). When heavy drinking or chronic drinking occurs, the acetaldehyde dehydrogenase enzyme cannot keep up with the Alcohol Dehydrogenase enzyme, and the pool of Acetaldehyde increases. This aldehyde has many toxic effects within biological systems (Figure \(\PageIndex{19}\):).
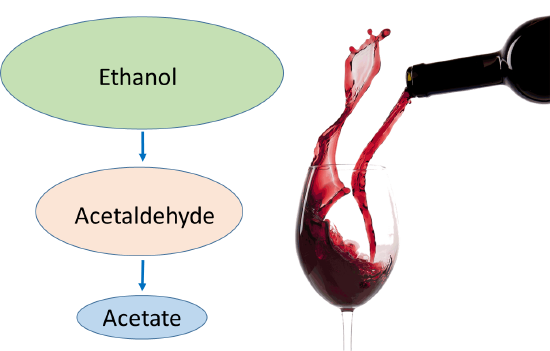
Figure \(\PageIndex{18}\): The Effects of Binge or Chronic Drinking on Ethanol Metabolites. Figure modified from Zakhari, S. (2013) Alcohol Research: Current Reviews 35(1):6-16
Acetaldehyde can form adducts with proteins. For example, acetaldehyde adducts on cytoskeletal components such as microtubulin lead to the swelling of hepatocytes (liver cells). If secreted from the cell, these protein adducts can also be recognized as foreign by the immune system and cause an autoimmune response, causing further inflammation and damage to the liver. Acetaldehyde also causes oxidative damage to lipids and DNA and can alter mitochondrial function. Overall, the liver is stressed and unhappy when too much ethanol is consumed.
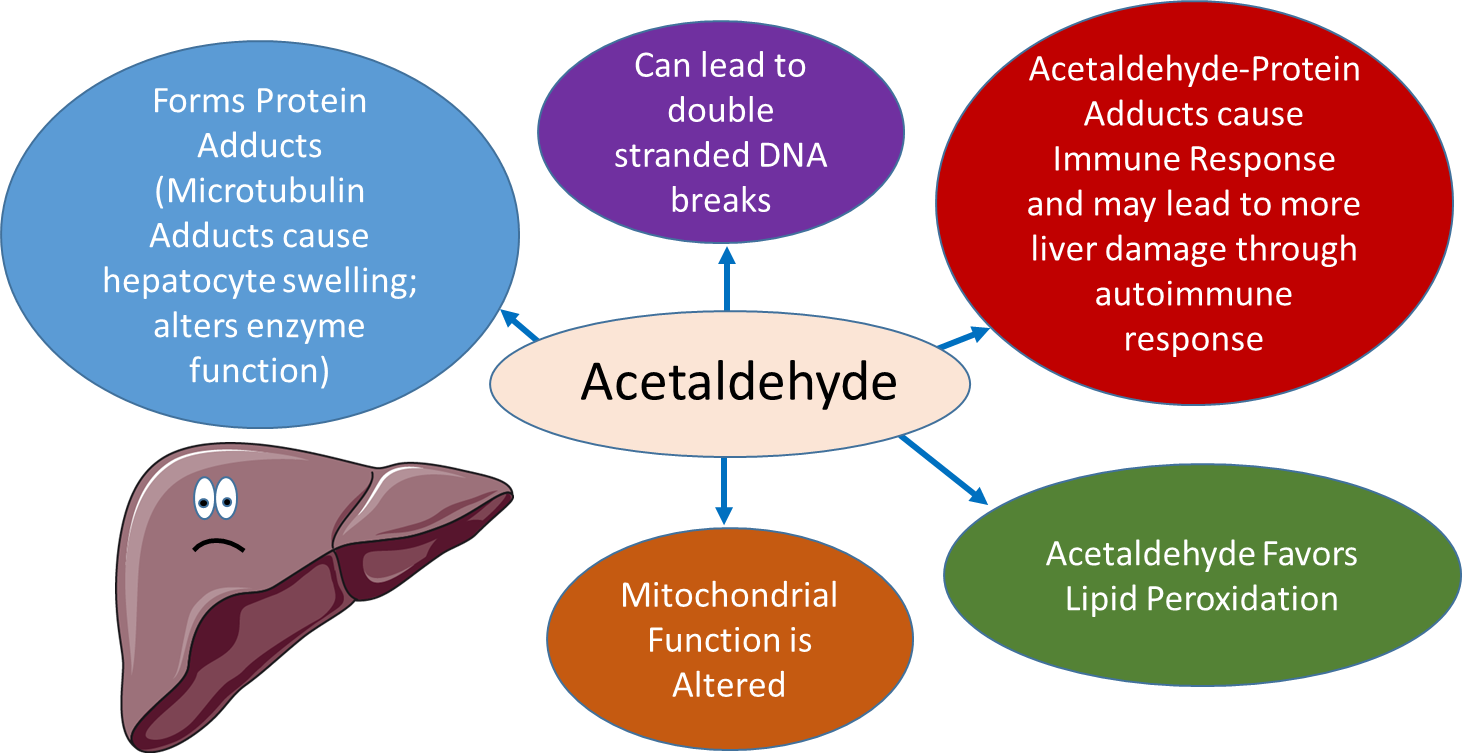
Figure \(\PageIndex{19}\): Acetaldehyde Toxicity. Figure modified from Servier Medical Art
Heavy chronic drinking can also lead to epigenetic modifications that alter protein expression patterns within the cell. Due to the oxidation of alcohol to acetate, alcohol metabolism also increases the NADH: NAD+ ratio, altering carbohydrate metabolism. Heavy ethanol consumption promotes the formation of reactive oxygen species and can also promote apoptosis. Overall, heavy drinking is extremely hard on liver function. It can lead progressively from the formation of a fatty liver to liver cirrhosis and increased risk of liver cancer. The risk for several other types of cancer is also heightened with heavy alcohol use.
Chronic heavy alcohol use can also alter gene expression, especially in the liver. Figure \(\PageIndex{20}\) is a schematic representation of DNA methylation, which converts cytosine to 5′methyl-cytosine via the actions of DNA methyltransferase (DNMT). DNA methylation typically occurs at cytosines that are followed by a guanine (i.e., CpG motifs). Within the liver, chronic heavy drinking reduces pools of S-adenosylmethionine (SAM) while increasing homocysteine and S-adenosylhomocysteine (SAH). SAH further inhibits DNA methyltransferases (DNMTs) by negative feedback inhibition, ultimately resulting in the global hypomethylation of DNA. This hypomethylation leads to the inappropriate expression of many genes, especially within the liver tissue.
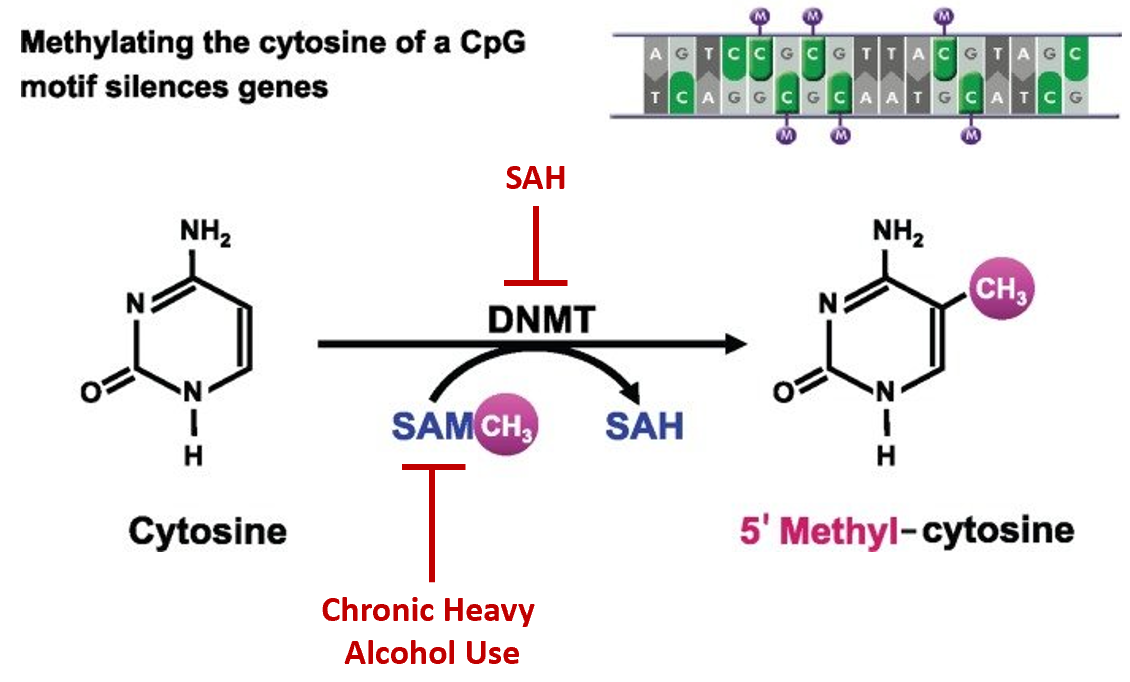
Figure \(\PageIndex{20}\): Effects of Chronic Alcohol Use on DNA Methylation. Figure modified from Zakhari, S. (2013) Alcohol Research: Current Reviews 35(1):6-16
Two upregulated genes due to this hypomethylation encode catalase and the p450 oxidoreductase, CYP2E1. Both of these enzymes are involved in oxidative pathways of alcohol metabolism that produce the toxic acetaldehyde intermediate (Figure \(\PageIndex{21}\)). Expression of both of these proteins becomes more prevalent in chronic alcohol consumption or when blood alcohol levels are high, as in cases of binge or heavy drinking. The activity of these enzymes can also lead to the formation of reactive oxygen species that contribute to global cellular damage (lipid peroxidation, DNA damage, protein damage, etc).
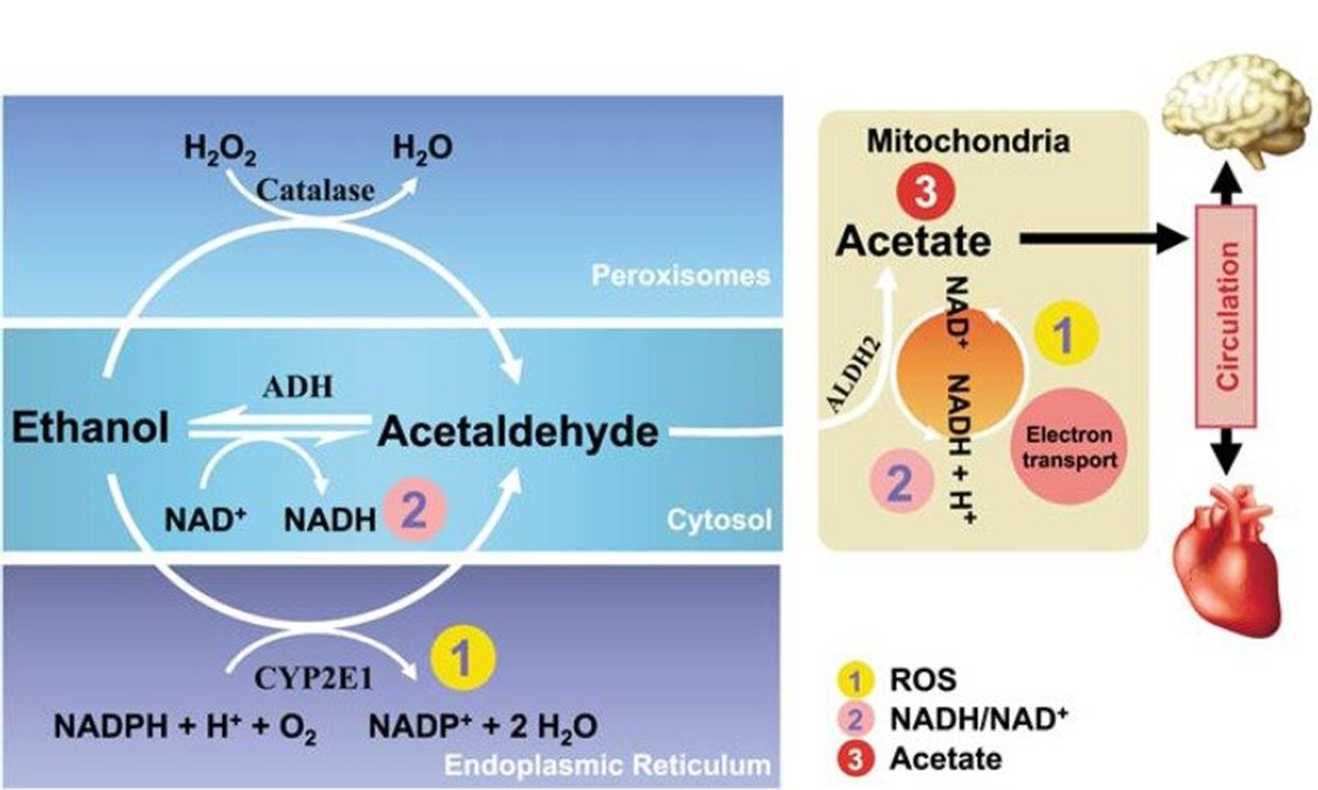
Figure \(\PageIndex{21}\): Oxidative Pathways of Alcohol Metabolism
Figure from Zakhari, S. (2013) Alcohol Research: Current Reviews 35(1):6-16
Gene regulation in other areas of the body is also affected in response to chronic heavy alcohol consumption. This is due to the production of acetate during the metabolic pathway of alcohol, which is released from the liver into the bloodstream. In other areas of the body, acetate is converted to acetyl-CoA by the enzyme Acetyl-Coenzyme A (acetyl-CoA) synthetase (AceCS). AceCS is activated by Sirtuin 1, also known as NAD-dependent protein deacetylase (SIRT1). Acetyl-CoA is used by histone acetyltransferase (HAT) to acetylate the lysine residues in histone proteins. Histone acetylation causes these proteins to release the bound DNA, allowing regions to be opened up for transcription (Figure \(\PageIndex{22}\)). Thus, higher levels of acetate promote histone acetylation and increased gene expression.
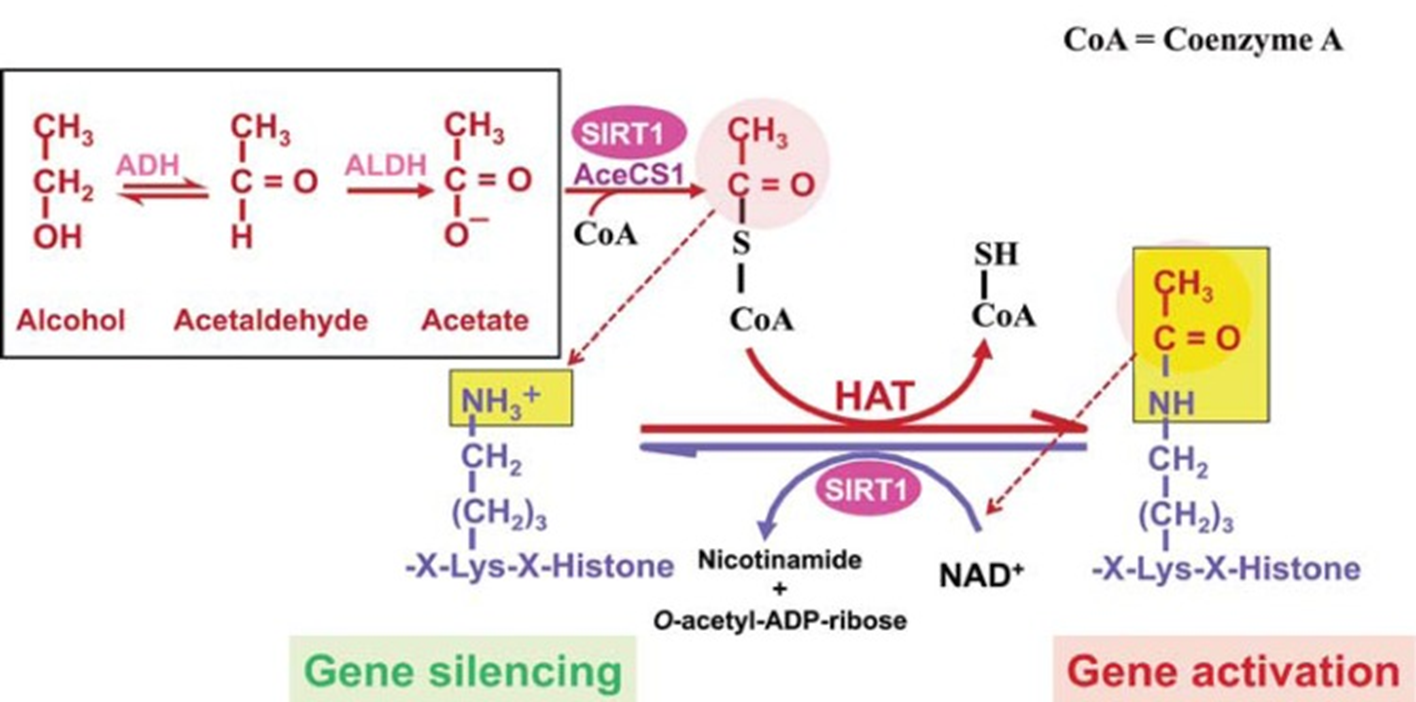
Figure \(\PageIndex{22}\): The Effects of Chronic Alcohol Consumption on Histone Acetylation. Note in the diagram shown, that SIRT1 also deacetylates histones, resulting in gene silencing. Thus, SIRT1 is a sensor that balances gene activation and silencing in the cell based on the cell’s energy status. Alcohol metabolism results in acetate formation, which is used in extrahepatic tissues to produce acetyl-CoA, upregulating histone acetylation within those tissues. NOTES: AceCS1 = Acetyl-CoA synthase 1; ADH = alcohol dehydrogenase; ALDH = Aldehyde dehydrogenase. Figure from Zakhari, S. (2013) Alcohol Research: Current Reviews 35(1):6-16
In addition to these effects, both ADH and ALDH utilize the cofactor nicotinamide adenine dinucleotide (NAD+), which is reduced to NADH; as a consequence, during ethanol oxidation, the ratio NADH/NAD+ is significantly increased, altering the cellular redox state and triggering several adverse effects related to alcohol consumption. Glycolysis and the Kreb Cycle are downregulated due to low NAD+ levels. This results in lower pyruvate levels and lower conversion of pyruvate to acetyl-CoA and also causes a decrease in gluconeogenesis (ie, there is not enough pyruvate to drive glucose production). Thus, the pyruvate that does form favors anaerobic conversion to lactate and can result in lactic acidosis or lowering of the blood pH levels. Low rates of gluconeogenesis can also contribute to hypoglycemia, which can be seen during binge drinking.
Hyperuricemia or an increase in blood levels of uric acid can also occur due, in part, to the increased production of ketone bodies and lactic acid. Both the ketone bodies and lactate can compete with uric acid for excretion into the urine within the kidney. The uric acid gets retained and heightens blood levels.
Ketone bodies typically form during periods of starvation when carbohydrate stores have been depleted. When the liver can no longer efficiently maintain blood glucose levels, it will break down fatty acids into ketone bodies and secrete these into the bloodstream (Figure \(\PageIndex{23}\)). Ketone bodies, such as acetoacetate, acetone, and D-beta-hydroxybutyrate (which isn’t a real ketone, but is still referred to as a ketone body), are released into the bloodstream to compensate for the reduced glucose levels. Brain, heart, and skeletal muscle tissue can utilize ketone bodies as an energy source. This is a good short-term solution to starvation. However, the formation of lactate and ketone bodies can severely reduce blood pH levels and induce a life-threatening state known as ketoacidosis. In addition to starvation, heavy alcohol consumption can induce ketogenesis inappropriately.
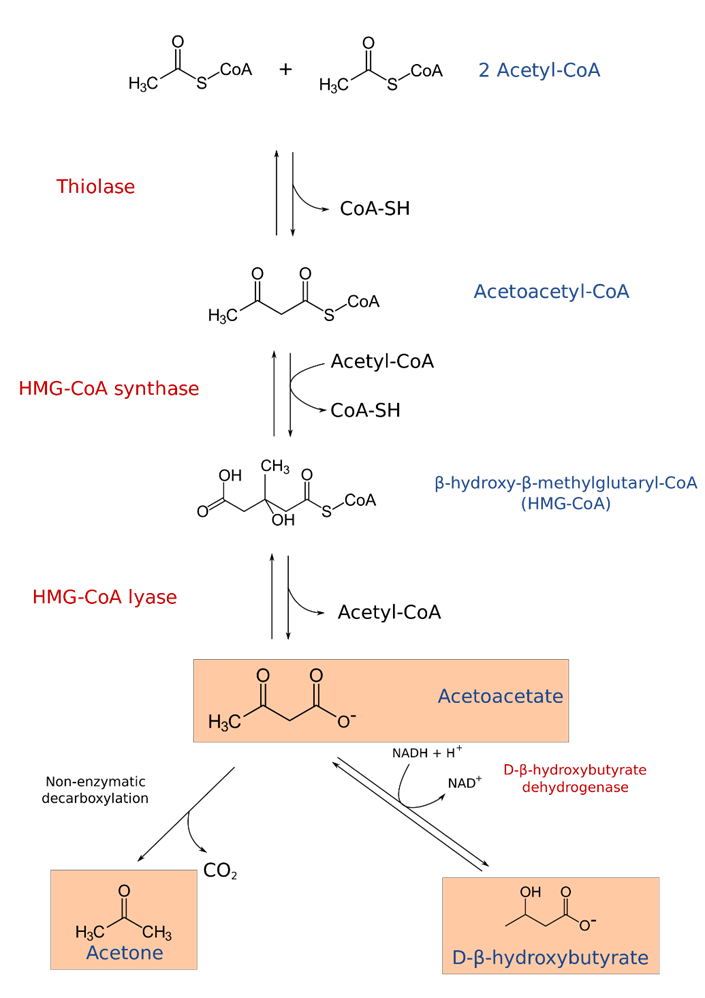
Figure \(\PageIndex{23}\): Formation of Ketone Bodies. Figure from Sav vas
As noted before, high levels of lactate and ketone bodies within the bloodstream can result in dehydration and reduced excretion of uric acid, leading to hyperuricemia. Over time, high uric acid levels can cause uric acid to precipitate, especially in joints, where it causes painful gout flare-ups (Figure \(\PageIndex{24}\)).
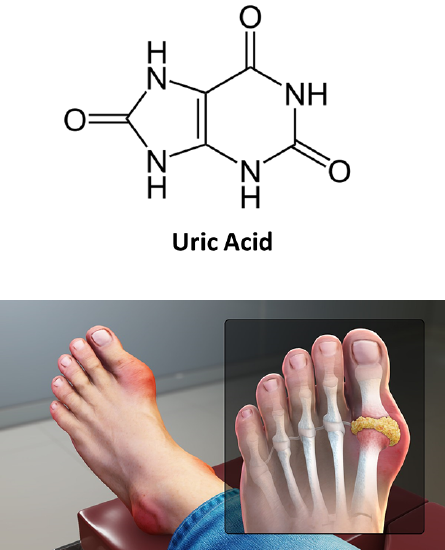
Figure \(\PageIndex{24}\): Hyperuricemia and Gout. Image from www.scientificanimations.com
According to the American Cancer Society, excessive alcohol use can also increase the risk for several different forms of cancer.
- Cancers of the mouth, throat, voice box, and esophagus: Alcohol use raises the risk of these cancers. Drinking and smoking together raise the risk of these cancers even more than drinking or smoking alone. This might be because alcohol can help harmful chemicals in tobacco get inside the cells that line the mouth, throat, and esophagus. Alcohol may also limit how these cells can repair damage to their DNA caused by the chemicals in tobacco.
- Liver cancer: Long-term alcohol use has been linked to an increased risk of liver cancer. Regular, heavy alcohol use can damage the liver, leading to inflammation and scarring. This might raise the risk of liver cancer.
- Colon and rectal cancer: Alcohol use has been linked with a higher risk of cancers of the colon and rectum. The evidence for this is generally stronger in men than in women, but studies have found the link in both sexes.
- Breast cancer: Even a few drinks a week are linked with an increased risk of breast cancer in women. This risk may be especially high in women who do not get enough folate (Vitamin B12) in their diet or through supplements. Alcohol can also raise estrogen levels in the body, which may explain some of the increased risks. Cutting back on alcohol may be an important way for many women to lower their risk of breast cancer.
So how does alcohol consumption contribute to increased risks of cancer? Alcohol may help other harmful chemicals, such as those in tobacco smoke, enter the cells lining the upper digestive tract more easily. This might explain why the combination of smoking and drinking is much more likely to cause cancers in the mouth or throat than smoking or drinking alone. In other cases, alcohol may slow the body’s ability to break down and get rid of some harmful chemicals. Alcohol might affect the body’s ability to absorb some nutrients, such as folate. Folate is a vitamin needed as a cofactor for enzymes involved in amino acid biosynthesis. Absorption of nutrients can be even worse in heavy drinkers, who often have low levels of folate. These low levels may play a role in the risk of some cancers, such as breast and colorectal cancer. Alcohol can raise the levels of estrogen, a hormone important in the growth and development of breast tissue. This could affect a woman’s risk of breast cancer. Too much alcohol can also add extra calories to the diet, which can contribute to weight gain in some people. Being overweight or obese is known to increase the risk of many types of cancer.
Overall, heavy ethanol consumption produces a wide spectrum of hepatic lesions (Figure \(\PageIndex{25}\)). Fatty liver (i.e., steatosis) is the earliest, most common response that develops in more than 90 percent of problem drinkers who consume 4 to 5 standard drinks per day. With continued drinking, alcoholic liver disease can proceed to liver inflammation (i.e., steatohepatitis), fibrosis, cirrhosis, and even liver cancer (i.e., hepatocellular carcinoma). Heavy drinking can also damage other organs, such as the pancreas and the brain, and can raise blood pressure. It also increases the risk of heart disease and stroke. In pregnant women, alcohol use, especially heavy drinking, may lead to birth defects or other problems with the fetus.
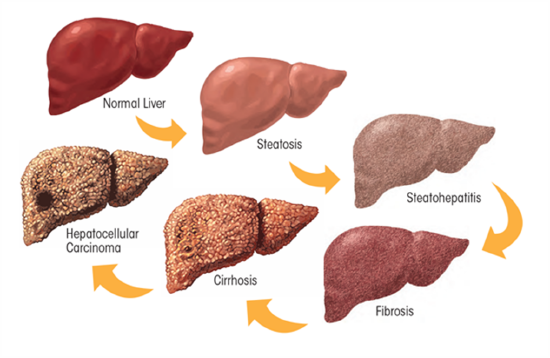
Figure \(\PageIndex{25}\): The Effects of Heavy Chronic Drinking on the Liver. Figure from Osna, N.A. et al (2017) Alcohol Research: Current Reviews 38(2)
In 1951, the FDA approved disulfiram for the treatment of alcoholism in the US (Figure \(\PageIndex{26}\)). Initially, the drug was prescribed in very high doses, often as high as 3,000 mg per day. The high doses led to reports of severe reactions to alcohol, some of which were fatal. It is an inhibitor of the ALDH-2, which will lead to an even higher increase in acetaldehyde concentration if alcohol is consumed. Thus, alcoholics who are taking this drug to quit drinking need to maintain sobriety. If they drink while taking this drug, they will become very ill due to the accumulation of acetaldehyde. Overall, the use of this drug supports abstinence, which is often hard to achieve realistically.
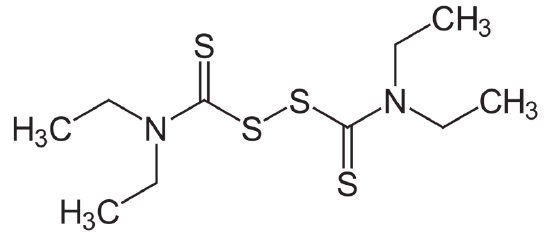
Figure \(\PageIndex{26}\): Disulfiram
At one time, it was thought that before prescribing disulfiram to people, patients should experience mixing the drug with alcohol in a supervised setting. Researchers felt it was important for the individuals to have full knowledge of what would happen if they mixed disulfiram and alcohol. This practice is no longer used, but every person must be educated on the reactions of combining alcohol with disulfiram before a prescription is written. Along with these effects, alcohol may contribute to cancer growth in other, unknown ways.
In other mammals, various ADHs are used to metabolize ethanol to acetaldehyde oxidatively (also toxic), which is converted to acetate by the enzyme aldehyde dehydrogenase, which catalyzes the following reaction:
CH3CHO + NAD+ + H2O → acetate + NADH + H+
They can also convert methanol to the very toxic and reactive formaldehyde, which makes methanol poisoning so dangerous, if not fatal. Mammalian enzymes are dimers of up to nine different monomers.
Summary
This chapter provides an in-depth exploration of fermentation as an anaerobic metabolic process that sustains ATP production when oxygen is scarce. It begins by emphasizing that glycolysis, which converts glucose into pyruvate while yielding 2 ATP and 2 NADH molecules, must be coupled with NADH oxidation to regenerate NAD⁺. Without this regeneration, glycolysis would eventually stall due to the limited pool of NAD⁺ available.
Two primary fermentative pathways are described in detail:
-
Lactate Fermentation:
In animal cells, particularly in muscle tissues under anaerobic conditions, pyruvate is reduced to lactate by lactate dehydrogenase (LDH). This reaction not only regenerates NAD⁺ but also allows glycolysis to continue during intense exercise or oxygen deficiency. The chapter reviews the enzyme’s structure, isozyme variations (e.g., LDH-1 in the heart, LDH-5 in muscle), and its reversible nature. Additionally, the Cori cycle is introduced as the mechanism by which lactate produced in muscles is transported to the liver, converted back to pyruvate, and ultimately used for gluconeogenesis, thereby linking peripheral anaerobic metabolism with hepatic aerobic processes. -
Ethanol Fermentation in Yeast:
Yeast and some other organisms utilize a two-step anaerobic process where pyruvate is first decarboxylated to acetaldehyde by thiamine pyrophosphate-dependent pyruvate decarboxylase, and then reduced to ethanol by alcohol dehydrogenase (ADH), regenerating NAD⁺ in the process. This pathway enables yeast to survive in oxygen-limited environments and underpins the industrial production of alcoholic beverages.
The chapter further extends its discussion to human alcohol metabolism, where ethanol is oxidized in the liver by cytosolic ADH to acetaldehyde, which is then further oxidized by mitochondrial acetaldehyde dehydrogenase (ALDH2) to acetate. This metabolism not only affects the cellular NADH/NAD⁺ ratio—impacting glycolysis, the Krebs cycle, and gluconeogenesis—but also has clinical significance. Excessive ethanol consumption leads to acetaldehyde accumulation, which is toxic and can cause liver damage, promote oxidative stress, and even contribute to cancer risk. The concept of hormesis is introduced to illustrate how moderate alcohol consumption may confer some cardioprotective effects, while chronic heavy drinking has severe detrimental consequences.
Lastly, the chapter touches on therapeutic strategies such as disulfiram, a drug that inhibits ALDH, thereby reinforcing abstinence by causing adverse reactions when alcohol is consumed.
Overall, the chapter weaves together the biochemical, physiological, and clinical aspects of fermentation and ethanol metabolism, underscoring their critical roles in energy production, metabolic regulation, and human health.