
6.5: RNA Editing
- Page ID
- 4837

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Occasionally researches encounter a gene with a sequence of nucleotides that does not match exactly that in its RNA product:
- messenger RNA (mRNA)
- ribosomal RNA (rRNA)
- transfer RNA (tRNA)
- microRNA (miRNA)
If the product is mRNA, some of the codons in the open reading frame (ORF) of the gene specify different amino acids from those in the protein translated from the mRNA of the gene.
The reason is RNA editing: the alteration of the sequence of nucleotides in the RNA
- after it has been transcribed from DNA but
- before it is translated into protein
RNA editing occurs by two distinct mechanisms:
- Substitution Editing: chemical alteration of individual nucleotides (the equivalent of point mutations).
These alterations are catalyzed by enzymes that recognize a specific target sequence of nucleotides (much like restriction enzymes):
- cytidine deaminases that convert a C in the RNA to uracil (U);
- adenosine deaminases that convert an A to inosine (I), which the ribosome translates as a G. Thus a CAG codon (for Gln) can be converted to a CGG codon (for Arg).
- Insertion/Deletion Editing: insertion or deletion of nucleotides in the RNA.
These alterations are mediated by guide RNA molecules that
- base-pair as best they can with the RNA to be edited and
- serve as a template for the addition (or removal) of nucleotides in the target
Substitution Editing
The human APOB gene
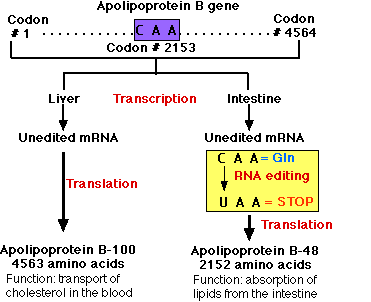
Humans have a single locus encoding the APOB gene.
- It contains 29 exons (separated by 28 introns).
- The exons contain a total of 4564 codons.
- Codon 2153 is CAA, which is a codon for the amino acid glutamine (Gln).
- The gene is expressed in cells of both the liver and the intestine.
- In both locations, transcription produces a pre-messenger RNA that must be spliced to produce the mRNA to be translated into protein.
- In the Liver. Here the process occurs normally producing apolipoprotein B-100 — a protein containing 4,563 amino acids — that is essential for the transport of cholesterol and other lipids in the blood.
- In the Intestine
- In the cells of the intestine, an additional step of pre-mRNA processing occurs: the chemical modification of the C nucleotide in Codon 2153 (CAA) into a U.
- This RNA editing changes the codon from one encoding the amino acid glutamine (Gln) to a STOP codon (UAA)
- The modification is catalyzed by the enzyme cytidine deaminase that
- recognizes the sequence of the RNA at that one place in the molecule and
- catalyzes the deamination of C thus forming U.
- Translation of the mRNA stops at codon #2153 forming apolipoprotein B-48 — a protein containing 2152 amino acids — that aids in the absorption of dietary lipids from the contents of the intestine.
DNA can also be edited. B cells express another cytidine deaminase (called activation-induced deaminase or AID) that is essential for both class switch recombination (CSR) and somatic hypermutation (SHM) of antibody genes. Humans with disabling mutations in the gene for this enzyme produce only IgM antibodies. However, here the enzyme is acting on DNA, not RNA. In attempting to repair the mismatch formed (dC•dG converted to dU•dG), the normal DNA repair machinery of the cell produces CSR or SHM as the situation warrants. (This process is also responsible for the occasional aberrant translocation of the heavy-chain gene segments to a proto-oncogene. The result is a B-cell cancer — a lymphoma or leukemia.)
Other examples of substitution editing
- Some mRNAs, tRNAs, and rRNAs in both the mitochondria and chloroplasts of plants;
- mRNAs encoding subunits of some receptors of neurotransmitters in the mammalian brain, e.g.,
- the AMPA receptor for Glu
- a serotonin receptor
- a tRNA in the mitochondria of the duckbill platypus
Insertion/Deletion Editing
The gene in mitochondria of Trypanosoma brucei
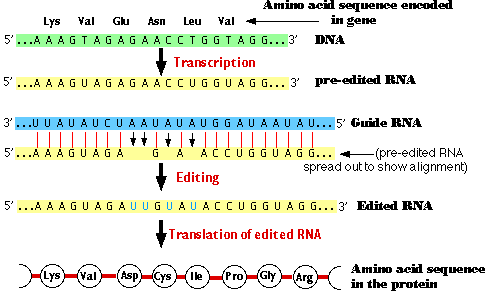
Several genes encoded in the mitochondrial DNA of this species (the cause of sleeping sickness in humans) encode transcripts that must be edited to make the mRNA molecules that will be translated into protein.
Editing requires a special class of RNA molecules called guide RNA (gRNA).
These small molecules have sequences that are complementary to the region around the site to be edited. The guide RNA base-pairs — as best it can — with this region. Note that in addition to the usual purine-pyrimidine pairing of C-G and A-U, G-U base-pairing can also occur.
Because of the lack of precise sequence complementarity, bulges occur either
- in the guide RNA where, usually, there are As not found in the transcript to be edited (as shown here) or
- in the transcript to be edited.
The bulges are eliminated by cutting the backbone of the shorter molecule and inserting complementary bases.
- In the first case (shown here) this produces insertions (here of Us)
- In the second case (not shown) this produces deletions.
Note that in the example shown here, the insertion of 4 nucleotides has created a frameshift so that the amino acids encoded downstream (after Val) in the edited RNA are entirely different from those specified by the gene itself.
Other examples of insertion/deletion editing
Insertion/deletion editing has also been found occur with
- mRNA, rRNA, and tRNA transcripts in the mitochondria of the slime mold Physarum polycephalum
- in measles virus transcripts
Why RNA Editing?
Good question. Some possibilities:
- So RNA editing appears to be here to stay. In fact, defects in RNA editing are associated with some human cancers as well as with amyotrophic lateral sclerosis (ALS — "Lou Gehrig's disease").