
13.2: Telomeres
- Page ID
- 5134

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Each eukaryotic chromosome consists of a single molecule of DNA associated with a variety of proteins. The DNA molecules in eukaryotic chromosomes are linear; i.e., have two ends. (This is in contrast to such bacterial chromosomes as that in E. coli that is a closed circle, i.e. has no ends.)
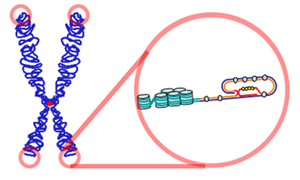
The DNA molecule of a typical chromosome contains a linear array of genes (encoding proteins and RNAs) interspersed with much noncoding DNA. Included in the noncoding DNA are long stretches that make up the centromere and long stretches at the ends of the chromosome, the telomeres. Telomeres are crucial to the life of the cell. They keep the ends of the various chromosomes in the cell from accidentally becoming attached to each other. The telomeres of humans consist of as many as 2000 repeats of the sequence 5' GGTTAG 3'
5'...GGTTAG GGTTAG GGTTAG GGTTAG GGTTAG GGTTAG..3'
3'...CCAATC CCAATC CCAATC CCAATC CCAATC CCAATC..5'
Replication of linear chromosomes presents a special problem
DNA polymerase can only synthesize a new strand of DNA as it moves along the template strand in the 3' –> 5' direction. This works fine for the 3' –> 5' strand of a chromosome as the DNA polymerase can move uninterruptedly from an origin of replication until it meets another bubble of replication or the end of the chromosome. However, synthesis using the 5' –> 3' strand as the template has to be discontinuous. When the replication fork opens sufficiently, DNA polymerase can begin to synthesize a section of complementary strand — called an Okazaki fragment — working in the opposite direction. Later, a DNA ligase ("DNA ligase I") stitches the Okazaki fragments together.
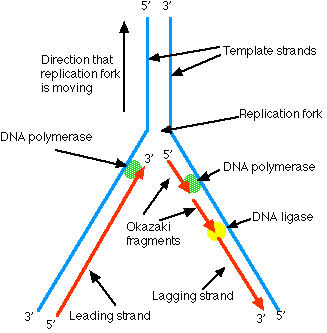
In Figure \(\PageIndex{3}\), the horizontal black arrows show the direction that the replication forks are moving. Wherever the replication fork of a strand is moving towards the 3' end, the newly-synthesized DNA (red) begins as Okazaki fragments (red dashes). This continues until close to the end of the chromosome. Then, as the replication fork nears the end of the DNA, there is no longer enough template to continue forming Okazaki fragments. So the 5' end of each newly-synthesized strand cannot be completed. Thus each of the daughter chromosomes will have a shortened telomere.
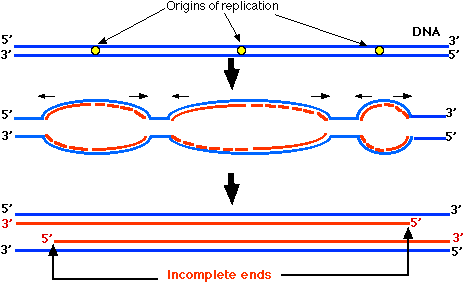
It is estimated that human telomeres lose about 100 base pairs from their telomeric DNA at each mitosis. This represents about 16 GGTTAG repeats. At this rate, after 125 mitotic divisions, the telomeres would be completely gone. Is this why normal somatic cells are limited in the number of mitotic divisions before they die out?
Telomeres and Cellular Aging
Telomeres are important so their steady shrinking with each mitosis might impose a finite life span on cells. This, in fact, is the case. Normal (non-cancerous) cells do not grow indefinitely when placed in culture. Cells removed from a newborn infant and placed in culture will go on to divide almost 100 times. Well before the end, however, their rate of mitosis declines (to less than once every two weeks). Were my cells to be cultured (I am 81 years old), they would manage only a couple of dozen mitoses before they ceased dividing and died out. This phenomenon is called replicative senescence. Could shrinkage of telomeres be a clock that determines the longevity of a cell lineage and thus is responsible for replicative senescence?
Evidence
Some cells do not undergo replicative senescence:
- the cells of the germline (the germplasm)
- unicellular eukaryotes like Tetrahymena thermophila
- stem cells, including "adult" stem cells and cancer stem cells.
It turns out that these cells are able to maintain the length of their telomeres. They do so with the aid of an enzyme telomerase.
Telomerase
Telomerase is an enzyme that adds telomere repeat sequences to the 3' end of DNA strands. By lengthening this strand, DNA polymerase is able to complete the synthesis of the "incomplete ends" of the opposite strand. Telomerase is a ribonucleoprotein. Its single snoRNA molecule — called TERC ("TElomere RNA Component") — provides an CCAAUC (in mammals) template to guide the insertion of GGTTAG. Its protein component — called TERT ("TElomere Reverse Transcriptase") — provides the catalytic action. Thus telomerase is a reverse transcriptase; synthesizing DNA from an RNA template.
Telomerase is generally found only in the cells of the germline, including embryonic stem (ES) cells; unicellular eukaryotes like Tetrahymena thermophila; and some, perhaps all, "adult" stem cells (including cancer stem cells) and "progenitor" cells enabling them to proliferate. When normal somatic cells are transformed in the laboratory with DNA expressing high levels of telomerase, they continue to divide by mitosis long after replicative senescence should have set in. And they do so without any further shortening of their telomeres. This remarkable demonstration (reported by Bodnar et. al. in the 16 January 1998 issue of Science) provides the most compelling evidence yet that telomerase and maintenance of telomere length are the key to cell immortality.
Telomere Deficiency Syndromes
A number of rare human diseases are caused by mutations in TERT or TERC or several other genes involved in telomere maintenance. The severity of the disease, the organs it affects, and the age of onset vary widely. But all are characterized by abnormally short telomeres. Patients with telomere deficiency are also at increased risk of developing cancer.
Telomerase and Cancer
The crucial feature that distinguishes a cancer from normal tissue is its ability to grow indefinitely. Most (85–90%) cancers express telomerase — at least in the population of cancer stem cells that divide uncontrollably causing the tumor to grow. Perhaps agents that prevent the expression of the gene for telomerase — or prevent the action of the enzyme — will provide a new class of weapons in the fight against cancer. But if telomerase activity — however brief — is essential for all cells, we had better be careful, and if lack of telomerase hastens replicative senescence, it may also hasten the aging of the tissues that depend on newly-formed cells for continued health — a tradeoff that may not be worth making.
Telomerase and Transplanted Cells
One approach to gene therapy it to remove cells from the patient, transform them with the gene for the product that the patient has been unable to synthesize, return them to the patient. One problem with this approach is that the cells — like all normal somatic cells — are mortal. After a series of mitotic divisions, they die out. That is the reason the children described in the link above required periodic fresh infusions of their transformed T cells.
What if their cells could be transformed not only with the therapeutic gene, but also with an active telomerase gene? This should give them an unlimited life span. But if cancer cells regain the ability to make telomerase, might not the reverse be true; that cells transformed with an active telomerase gene might become cancerous? Perhaps not. The cells described by Bodnar et. al. in the 16 January 1998 issue of Science have continued to grow in culture and have been subjected to a number of tests to see if they have acquired any properties of cancer cells in culture.
The results are encouraging. While these cells continue to divide indefinitely as cancer cells do,
- They still show contact inhibition as normal cells do when grown in culture.
- They do not grow into tumors when injected into immunodeficient mice (as cancer cells do).
- They are still fussy about their diet — unable to grow on the simple media that supports cancer cells in culture.
- They still retain a normal karyotype; something that cancer cells seldom do.
However, studies with whole animals — transgenic mice that express abnormally high levels of TERT — reveal that they do suffer an elevated incidence of cancer.
The now-famous sheep Dolly was cloned using a nucleus taken from an adult sheep cell that had been growing in culture. The cell donor was 6 years old, and its cells had been growing in culture for several weeks. What about Dolly's telomeres? Analysis of telomere length in Dolly's cells reveals that they were only 80% as long as in a normal one-year-old sheep. Not surprising, since the nucleus that created Dolly had been deprived of telomerase for many generations. Two other sheep — cloned from embryonic, not adult, cells — also had shortened telomeres although not as short as Dolly's. Perhaps the length of time the cells spent in culture before they were used accounts for this.

Dolly. (Cc BY-SA 2.0 Toni Barros).
Does this mean that Dolly is doomed to a shortened life? She seemed healthy at first and even had babies of her own. But medical problems — probably unrelated to her telomeres — ended with her being euthanized at a relatively young age. But her short telomeres do add another question to the debate about cloning mammals from adult cells.