
15.6.1.7: Sex Hormones
- Page ID
- 5477

The ovaries of sexually-mature females secrete both a mixture of estrogens (of which 17 β-estradiol is the most abundant and most potent) and progesterone.
Estrogens
Estrogens are steroids and are primarily responsible for the conversion of girls into sexually-mature women including:
- development of breasts
- further development of the uterus and vagina
- broadening of the pelvis
- growth of pubic and axillary hair
- increase in adipose (fat) tissue
- participate in the monthly preparation of the body for a possible pregnancy
- participate in pregnancy if it occurs
Estrogens also have non-reproductive effects. For example, they antagonize the effects of the parathyroid hormone, minimizing the loss of calcium from bones and thus helping to keep bones strong. They also promote blood clotting.
Progesterone
Progesterone is also a steroid. It has many effects in the body, some having nothing to do with sex and reproduction. Here we shall focus on the role of progesterone in the menstrual cycle and pregnancy.
How estrogens and progesterone achieve their effects
Steroids like estrogens and progesterone are small, hydrophobic molecules that are transported in the blood bound to a serum globulin.
- In "target" cells, i.e., cells that change their gene expression in response to the hormone, they bind to receptor proteins located in the cytoplasm and/or nucleus.
- The hormone-receptor complex enters the nucleus (if it formed in the cytoplasm) and binds to specific sequences of DNA, called the estrogen (or progesterone) response elements.
- Response elements are located in the promoters of genes.
- The hormone-receptor complex acts as a transcription factor (often recruiting other transcription factors to help) which turns on (sometimes off) transcription of those genes.
- Gene expression in the cell produces the response.
Some "target" cells also have other types of estrogen and progesterone receptors that are embedded in a membrane (endoplasmic reticulum and plasma membrane respectively). Binding of the hormone to them produces more rapid effects than those of the nuclear receptors. For example, human sperm have receptors that within a second of being exposed to progesterone activate the sperm to increased motility.
Regulation of Estrogen and Progesterone
The synthesis and secretion of estrogens is stimulated by follicle-stimulating hormone (FSH), which is, in turn, controlled by the hypothalamic gonadotropin releasing hormone (GnRH).
| Hypothalamus | → | GnRH | → | Pituitary | → | FSH | → | Follicle | → | Estrogens |
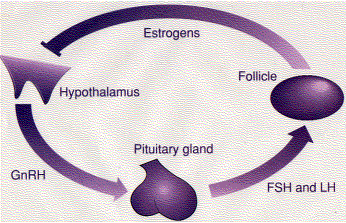
High levels of estrogens suppress the release of GnRH (bar) providing a negative-feedback control of hormone levels.
It works like this: Secretion of GnRH depends on certain neurons in the hypothalamus which express a gene (KISS-1) encoding a protein of 145 amino acids. From this are cut several short peptides collectively called kisspeptin. These are secreted and bind to G-protein-coupled receptors on the surface of the GnRH neurons stimulating them to release GnRH. However, high levels of estrogen (or progesterone or testosterone) inhibit the secretion of kisspeptin and suppress further production of those hormones.
Progesterone production is stimulated by luteinizing hormone (LH), which is also stimulated by GnRH.
| Hypothalamus | → | GnRH | → | Pituitary | → | LH | → | Corpus luteum | → | Progesterone |
Elevated levels of progesterone control themselves by the same negative feedback loop used by estrogen (and testosterone).
The Menstrual Cycle
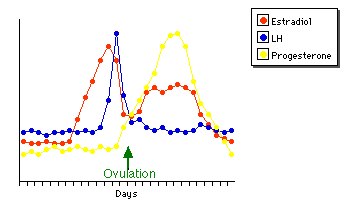
About every 28 days, some blood and other products of the disintegration of the inner lining of the uterus (the endometrium) are discharged from the uterus, a process called menstruation. During this time a new follicle begins to develop in one of the ovaries. After menstruation ceases, the follicle continues to develop, secreting an increasing amount of estrogen as it does so.
- The rising level of estrogen causes the endometrium to become thicker and more richly supplied with blood vessels and glands.
- A rising level of LH causes the developing egg within the follicle to complete the first meiotic division (meiosis I), forming a secondary oocyte.
- After about two weeks, there is a sudden surge in the production of LH.
- This surge in LH triggers ovulation: the release of the secondary oocyte into the fallopian tube.
- Under the continued influence of LH, the now-empty follicle develops into a corpus luteum (hence the name luteinizing hormone for LH).
- Stimulated by LH, the corpus luteum secretes progesterone which
- continues the preparation of the endometrium for a possible pregnancy
- inhibits the contraction of the uterus
- inhibits the development of a new follicle
- If fertilization does not occur (which is usually the case)
- the rising level of progesterone inhibits the release of GnRH which, in turn, inhibits further production of progesterone
- As the progesterone level drops
- the corpus luteum begins to degenerate
- the endometrium begins to break down, its cells committing programmed cell death
- the inhibition of uterine contraction is lifted
- the bleeding and cramps of menstruation begin.
Fertilization
Fertilization of the egg is also influenced by progesterone. Sperm swim towards the egg by chemotaxis following a gradient of progesterone secreted by cells surrounding the egg. Progesterone opens CatSper ("cation sperm") channels in the plasma membrane surrounding the anterior portion of the sperm tail. This allows an influx of Ca2+ ions which causes the flagellum to beat more rapidly and vigorously.
Pregnancy
As the fertilized egg passes down the fallopian tube, it undergoes its first mitotic divisions. By the end of the week, the developing embryo has become a hollow ball of cells called a blastocyst. At this time, the blastocyst reaches the uterus and embeds itself in the endometrium, a process called implantation. With implantation, pregnancy is established.
The blastocyst has two parts:
- the inner cell mass, which will become the baby
- the trophoblast, which will
- develop into the placenta and umbilical cord
- begin to secrete human chorionic gonadotropin (HCG).
HCG is a glycoprotein. It is a heterodimer of the same alpha subunit (of 92 amino acids) used by TSH, FSH, and LH and a unique beta subunit (of 145 amino acids). HCG behaves much like FSH and LH with one crucial exception: it is NOT inhibited by a rising level of progesterone. Thus HCG prevents the deterioration of the corpus luteum at the end of the fourth week and enables pregnancy to continue beyond the end of the normal menstrual cycle. Because only the implanted trophoblast makes HCG, its early appearance in the urine of pregnant women provides the basis for the most widely used test for pregnancy (which can provide a positive signal even before menstruation would have otherwise begun). As pregnancy continues, the placenta becomes a major source of progesterone, and its presence is essential to maintain pregnancy. Mothers at risk of giving birth too soon can be given a synthetic progestin to help them retain the fetus until it is full-term.
Birth
Toward the end of pregnancy,
- The placenta releases large amounts of CRH which stimulates the pituitary glands of both mother and her fetus to secrete.
- ACTH, which acts on their adrenal glands causing them to release the estrogen precursor dehydroepiandrosterone sulfate (DHEAS).
- This is converted into estrogen by the placenta.
- The rising level of estrogen causes the smooth muscle cells of the uterus to
- synthesize connexins and form gap junctions. Gap junctions connect the cells electrically so that they contract together as labor begins.
- express receptors for oxytocin.
- Oxytocin is secreted by the posterior lobe of the pituitary as well as by the uterus.
- Prostaglandins are synthesized in the placenta and uterus.
- The normal inhibition of uterine contraction by progesterone is turned off by several mechanisms while
- both oxytocin and prostaglandins cause the uterus to contract and labor begins.
Three or four days after the baby is born, the breasts begin to secrete milk. Milk synthesis is stimulated by the pituitary hormone prolactin (PRL), and its release from the breasts is stimulated by oxytocin. Milk contains an inhibitory peptide. If the breasts are not fully emptied, the peptide accumulates and inhibits milk production. This autocrine action thus matches supply with demand.
Other Hormones
- Relaxin
As the time of birth approaches in some animals (e.g., pigs, rats), this polypeptide has been found to:- relax the pubic ligaments
- soften and enlarge the opening to the cervix
Relaxin is found in pregnant humans but at higher levels early in pregnancy than close to the time of birth. Relaxin promotes angiogenesis, and in humans it probably plays a more important role in the development of the interface between the uterus and the placenta that it does in the birth process.
- Activins, Inhibins, Follistatin.
These proteins are synthesized within the follicle. Activins and inhibins bind to follistatin. Activins increase the action of FSH; inhibins, as their name suggests, inhibit it. How important they are in humans remains to be seen. However the important role that activin and follistatin play in the embryonic development of vertebrates justifies mentioning them here.
Oral contraceptives - the "pill"
The feedback inhibition of GnRH secretion by estrogens and progesterone provides the basis for the most widely-used form of contraception. Dozens of different formulations of synthetic estrogens or progestins (progesterone relatives or both are available. Their inhibition of GnRH prevents the mid-cycle surge of LH and ovulation. Hence there is no egg to be fertilized. Usually the preparation is taken for about three weeks and then stopped long enough for normal menstruation to occur. The main side-effects of the pill stem from an increased tendency for blood clots to form (estrogen enhances clotting of the blood).
RU-486
RU-486 (also known as mifepristone) is a synthetic steroid related to progesterone. Unlike the synthetic progestins used in oral contraceptives that mimic the actions of progesterone, RU-486 is a progesterone antagonist; that is, it blocks the action of progesterone. It does this by binding more tightly to the progesterone receptor than progesterone itself but without the normal biological effects:
- The RU-486/receptor complex is not active as a transcription factor.
- Thus genes that are turned on by progesterone are turned off by RU-486.
- The proteins needed to establish and maintain pregnancy are no longer synthesized.
- The endometrium breaks down.
- The embryo detaches from it and can no longer make chorionic gonadotropin (HCG).
- Consequently the corpus luteum ceases its production of progesterone.
- The inhibition on uterine contraction is lifted.
- Soon the embryo and the breakdown products of the endometrium are expelled.
These properties of RU-486 have caused it to be used to induce abortion of an unwanted fetus. In practice, the physician assists the process by giving a synthetic prostaglandin (e.g., misoprostol [Cytotec®]) 36–48 hours after giving the dose of RU-486. Use of RU-486 is generally limited to the first seven weeks of pregnancy. RU-486 has been used for many years in some countries. However, the controversies surrounding abortion in the United States kept it from being authorized for use here until September 2000.
Menopause
The menstrual cycle continues for many years. But eventually, usually between 42 and 52 years of age, the follicles become less responsive to FSH and LH. They begin to secrete less estrogen. Ovulation and menstruation become irregular and finally cease. This cessation is called menopause.
With levels of estrogen now running one-tenth or less of what they had been, the hypothalamus is released from their inhibitory influence (bar). As a result it now stimulates the pituitary to increased activity. The concentrations of FSH and LH in the blood rise to ten or more times their former values. These elevated levels may cause a variety of unpleasant physical and emotional symptoms.
Hormone Replacement Therapy (HRT)
Many menopausal women elect to take a combination of estrogen and progesterone after they cease to make their own. The benefits are (1) reduction in the unpleasant symptoms of the menopause and (2) a reduction in the loss of calcium from bones and thus a reduction in osteoporosis and the fractures that accompany it. It was also believed that HRT reduced the risk of cardiovascular disease. However, a recent study of 16,000 menopausal women was stopped 3 years early when it was found that, in fact, HRT increased (albeit only slightly) not decreased the incidence of cardiovascular disease. Perhaps synthetic selective estrogen response modulators or SERMs (raloxifene is an example) will provide the protective effects without the harmful ones.
Environmental estrogens
Some substances that find their way into the environment, such as
- DDE, a breakdown product of the once widely-used insecticide DDT
- DDT itself — still used in some countries (e.g., Mexico)
- PCBs, chemicals once used in a wide variety of industrial applications
can bind to the estrogen (and androgen) receptors and mimic (weakly) the effects of the hormone. This has created anxiety that they may be responsible for harmful effects such as cancer and low sperm counts.
However, there is as yet little evidence to support these worries. No epidemiological relationship has been found between the incidence of breast cancer and the levels of these compounds in the body. As for laboratory studies that found a synergistic effect of two of these substances on receptor binding (findings that created the great alarm), these have not been replicated in other laboratories, and the authors of the original report have since withdrawn it as invalid.
Males
The principal androgen (male sex hormone) is testosterone. This steroid is manufactured by the interstitial (Leydig) cells of the testes. Secretion of testosterone increases sharply at puberty and is responsible for the development of the so-called secondary sexual characteristics (e.g., beard) of men.
Testosterone is also essential for the production of sperm. Production of testosterone is controlled by the release of luteinizing hormone (LH) from the anterior lobe of the pituitary gland, which is in turn controlled by the release of GnRH from the hypothalamus. LH is also called interstitial cell stimulating hormone (ICSH).
| Hypothalamus | → | GnRH | → | Pituitary | → | LH | → | Testes | → | Testosterone |
The level of testosterone is under negative-feedback control: a rising level of testosterone suppresses the release of GnRH from the hypothalamus. This is exactly parallel to the control of estrogen secretion in females.
In mice, osteocalcin, a hormone secreted by osteoblasts of the bone, stimulates the synthesis of testosterone by Leydig cells even more powerfully than LH. Whether this effect occurs in humans remains to be seen.
In 1994, a man was described who was homozygous for a mutation in the gene encoding the estrogen receptor. A single nonsense mutation had converted a codon (CGA) for arginine early in the protein into a STOP codon (TGA). Thus no complete estrogen receptor could be synthesized.
This man was extra tall, had osteoporosis and "knock-knees", but was otherwise well. His genetic defect confirms the important role that estrogen has in both sexes for normal bone development. It is not known whether this man was fertile or not. However, mutations in their estrogen receptor gene have been found in other men who are sterile, and male mice whose estrogen receptor gene has been "knocked out" are sterile.
Another function of estrogen in males. The accumulation of fat in the abdomen, so characteristic of aging males (including yours truly), is caused by declining levels of estrogen.
Anabolic steroids
A number of synthetic androgens are used for therapeutic purposes. These drugs promote an increase in muscle size with resulting increases in strength and speed. This has made them popular with some athletes, e.g., weight lifters, cyclists, runners, swimmers, professional football players. Usually these athletes (females as well as males) take doses far greater than those used in standard therapy. Such illicit use carries dangers (besides being banned from an event because of a positive drug test): acne, a decrease in libido, and — in males — testicle size and sperm counts to name a few.
Genetic abnormalities of gonadal function
Many things can go wrong with sexual development in both males and females, fortunately rarely. Let's look at a few that clearly result from the inheritance of single-gene mutations.
- Inherited mutations in both copies of the gene encoding the GnRH receptor result in failure to develop at puberty.
- Mutations in the gene encoding the LH receptor prevent normal sexual development in both sexes.
- Mutations in the gene encoding the FSH receptor block development of the gonads in both males and females.
- Mutations in any of the genes encoding the enzymes for synthesis and metabolism of testosterone interfere with normal sexual function in males.
- A similar spectrum of disorders in males can be caused by mutations in the genes encoding the androgen receptor.
Contributors and Attributions
John W. Kimball. This content is distributed under a Creative Commons Attribution 3.0 Unported (CC BY 3.0) license and made possible by funding from The Saylor Foundation.