
2.6: Community Ecology
- Page ID
- 164553

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Introduction to Species Interactions
Populations rarely, if ever, live in isolation from populations of other species. All populations occupying the same habitat form a community (populations of multiple species inhabiting a specific area at the same time). The number of species occupying the same habitat and their relative abundance is known as species diversity. Areas with low diversity, such as the glaciers of Antarctica, still contain a wide variety of living things, whereas the diversity of tropical rainforests is so great that it cannot be counted. Ecology is studied at the community level to understand how species interact with each other and compete for the same resources.
The interactions among populations of different species play a major role in regulating population growth and abundance. A species interaction is the effect that a pair of organisms living together in a community have on each other. Interactions range from mutualism, which benefits both species involved, to competition, which harms both species involved (Wootton and Emmerson 2005). Interactions can be indirect, through intermediaries such as shared resources or common enemies. All of these interactions can be organized by the effects the species have on each other (Figure \(\PageIndex{1}\)).
Species interactions may be short-term, like pollination and predation, or long-term; both often strongly influence the evolution of the species involved. Short-term interactions are short-lived in terms of the duration of a single interaction: a predator kills and eats a prey; a pollinator transfers pollen from one flower to another; but they are extremely durable in terms of their influence on the evolution of both partners. As a result, the partners coevolve (Bengtson 2002, Lunau 2004).

Figure \(\PageIndex{1}\): Summary of species interactions, showing how each species is impacted by the interaction. Produced by N. Gownaris.

Figure \(\PageIndex{2}\): The black walnut secretes a chemical from its roots that harms neighboring plants, an example of competitive antagonism.

Figure \(\PageIndex{3}\): Predation is a short-term interaction, in which the predator, here an osprey, kills and eats its prey.
Competition
Competition can be defined as an interaction between organisms or species, in which the fitness of one is lowered by the presence of another. Competition is often for a resource such as food, water, or territory in limited supply, or for access to females for reproduction (Begon et al. 1996). Competition among members of the same species is known as intraspecific competition, while competition between individuals of different species is known as interspecific competition.

Figure \(\PageIndex{4}\): Direct intraspecific competition for mates between two male red deer.
The competitive exclusion principle postulates that two species which compete for the same limited resource cannot coexist at constant population values. When one species has even the slightest advantage over another, the one with the advantage will dominate in the long term. This leads either to the extinction of the weaker competitor or to an evolutionary or behavioral shift toward a different ecological niche. The principle has been paraphrased in the maxim "complete competitors cannot coexist" (Hardin, 1960).
Georgy Gause formulated the law of competitive exclusion based on laboratory competition experiments using two species of Paramecium, P. aurelia and P. caudatum. The conditions were to add fresh water every day and input a constant flow of food. Although P. caudatum initially dominated, P. aurelia recovered and subsequently drove P. caudatum extinct via exploitative resource competition. However, Gause was able to let the P. caudatum survive by differing the environmental parameters (food, water). Thus, Gause's law is valid only if the ecological factors are constant.
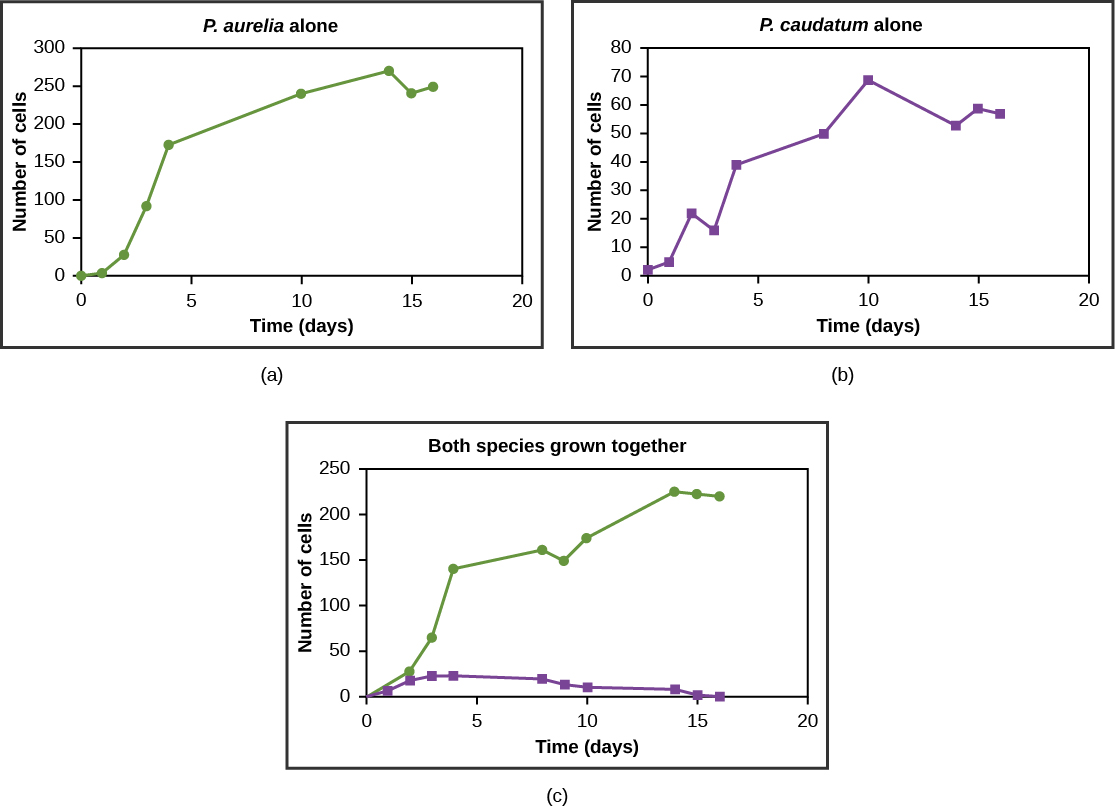
Figure \(\PageIndex{5}\): Paramecium aurelia and Paramecium caudatum grow well individually, but when they compete for the same resources, P. aurelia outcompetes P. caudatum.
However, competitive exclusion is rarely observed in natural ecosystems and many biological communities appear to violate Gause's law. The best-known example is the so-called "paradox of the plankton" (Hutchinson, 1960). All plankton species live on a very limited number of resources, primarily solar energy and minerals dissolved in the water. According to the competitive exclusion principle, only a small number of plankton species should be able to coexist on these resources. Nevertheless, large numbers of plankton species coexist within small regions of open sea.
If a species is particularly effective at co-opting resources to its own benefit, it may displace other species, a phenomenon known as competitive displacement (or in extreme cases, competitive exclusion). This affects the presence and relative abundance of species in the community. For example, sugar maple (Acer saccharum) is a highly competitive tree in hardwood forests of eastern Canada. Where environmental conditions are well suited for this species, it can dominate mature stands. If large sugar maple trees are removed from a stand, perhaps by a selective timber harvest, other tree species (as well as small sugar maples) will benefit from the reduced competition and will grow more vigorously.
This exclusion may be avoided if a population evolves to make use of a different resource, a different area of the habitat, or feeds during a different time of day, called resource partitioning. The two organisms are then said to occupy different microniches. These organisms coexist by minimizing direct competition.

Figure \(\PageIndex{6}\): 1) A smaller (yellow) species of bird forages across the whole tree. 2) A larger (red) species competes for resources. 3) Red dominates in the middle for the more abundant resources. Yellow shifts to a new niche, avoiding competition.
Niche differentiation (also known as niche separation and niche partitioning) refers to the process by which competing species use the environment differently in a way that helps them to coexist. When two species differentiate their niches, they tend to compete less strongly, and are thus more likely to coexist. Species can differentiate their niches in many ways, such as by consuming different foods, or using different areas of the environment. As an example of niche partitioning, several anole lizards in the Caribbean islands share common diets—mainly insects. They avoid competition by occupying different physical locations. Although these lizards might occupy different locations, some species can be found inhabiting the same range, with up to 15 in certain areas. For example, some live on the ground while others are arboreal. Species who live in different areas compete less for food and other resources, which minimizes competition between species. However, species who live in similar areas typically compete with each other (Pacala, 1985).
Resource Partitioning is the phenomenon where two or more species divide out resources like food, space, resting sites etc. to coexist. For example, some lizard species appear to coexist because they consume insects of differing sizes (Caldwell & Vitt, 1999). Alternatively, species can coexist on the same resources if each species is limited by different resources, or differently able to capture resources. Different types of phytoplankton can coexist when different species are differently limited by nitrogen, phosphorus, silicon, and light (Grover, 1997). In the Galapagos Islands, finches with small beaks are more able to consume small seeds, and finches with large beaks are more able to consume large seeds. If a species' density declines, then the food it most depends on will become more abundant (since there are so few individuals to consume it). As a result, the remaining individuals will experience less competition for food. Although "resource" generally refers to food, species can partition other non-consumable objects, such as parts of the habitat. For example, warblers are thought to coexist because they nest in different parts of trees (MacArthur, 1958). Species can also partition habitat in a way that gives them access to different types of resources. As previously stated, anole lizards appear to coexist because each uses different parts of the forests as perch locations (Grover, 1997). This likely gives them access to different species of insects.

Figure \(\PageIndex{7}\): Several species of Galapagos finches display character displacement. Each closely related species differs in beak size and beak depth, allowing them to coexist in the same region since each species eats a different type of seed: the seed best fit for its unique beak. The finches with the deeper, stronger beaks consume large, tough seeds, while the finches with smaller beaks consume the smaller, softer seeds.
Character displacement is the phenomenon where differences among similar species whose distributions overlap geographically are accentuated in regions where the species co-occur, but are minimized or lost where the species' distributions do not overlap. This pattern results from evolutionary change driven by biological competition among species for a limited resource (e.g. food). The rationale for character displacement stems from the competitive exclusion principle, which contends that to coexist in a stable environment two competing species must differ in their respective ecological niche; without differentiation, one species will eliminate or exclude the other through competition.
For example, Darwin's finches can be found alone or together on the Galapagos Islands. Both species' populations actually have more individuals with intermediate-sized beaks when they live on islands without the other species present. However, when both species are present on the same island, competition is intense between individuals that have intermediate-sized beaks of both species because they all require intermediate sized seeds. Consequently, individuals with small and large beaks have greater survival and reproduction on these islands than individuals with intermediate-sized beaks. Different finch species can coexist if they have traits—for instance, beak size—that allow them to specialize on particular resources. When Geospiza fortis and Geospiza fuliginosa are present on the same island, G. fuliginosa tends to evolve a small beak and G. fortis a large beak. The observation that competing species' traits are more different when they live in the same area than when competing species live in different areas is called character displacement. For the two finch species, beak size was displaced: beaks became smaller in one species and larger in the other species.
Mutualism
A mutualism occurs when two species benefit from their interaction. For example, termites have a mutualistic relationship with protists that live in the insect’s gut (Figure \(\PageIndex{8}\)a). The termite benefits from the ability of the protists to digest cellulose. However, the protists are able to digest cellulose only because of the presence of symbiotic bacteria within their cells that produce the cellulase enzyme. The termite itself cannot do this: without the protozoa, it would not be able to obtain energy from its food (cellulose from the wood it chews and eats). The protozoa benefit by having a protective environment and a constant supply of food from the wood chewing actions of the termite. In turn, the protists benefit from the enzymes provided by their bacterial endosymbionts, while the bacteria benefit from a doubly protective environment and a constant source of nutrients from two hosts.
Lichen are a mutualistic relationship between a fungus and photosynthetic algae or cyanobacteria (Figure \(\PageIndex{8}\)b). The glucose produced by the algae provides nourishment for both organisms, whereas the physical structure of the lichen protects the algae from the elements and makes certain nutrients in the atmosphere more available to the algae. The algae of lichens can live independently given the right environment, but many of the fungal partners are unable to live on their own.

Figure \(\PageIndex{8}\): (a) Termites form a mutualistic relationship with symbiotic protozoa in their guts, which allow both organisms to obtain energy from the cellulose the termite consumes. (b) Lichen is a fungus that has symbiotic photosynthetic algae living in close association. (credit a: modification of work by Scott Bauer, USDA; credit b: modification of work by Cory Zanker)
Another set of mutualisms are pollinators, such as bees, butterflies, and hummingbirds. Polinators benefit because they eat the collect pollen and/or nectar that they collect from flowers. Plants and pollinators are often coevolved (Ehrlich and Raven 1964; Pollan 2001; Lunau 2004). Insect-pollinated flowers have bright colors, patterns, scent, nectar, and sticky pollen to attract insects, guide them to pick up and deposit pollen, and reward pollinators. Conversely, pollinator insects like bees are adapted to detect flowers by color, pattern, and scent, to collect pollen (such as with bristles shaped to form pollen baskets on their hind legs), and nectar.

Figure \(\PageIndex{9}\): Hummingbird hawkmoth drinking from Dianthus.
Seed dispersal is the movement, spread or transport of seeds. Plants have limited mobility and rely upon a variety of dispersal vectors to transport their propagules, including both abiotic vectors such as the wind and living (biotic) vectors like birds (Lim ad Burns 2021) patterns of seed dispersal are determined in large part by the dispersal mechanism and this has important implications for the demographic and genetic structure of plant populations, as well as migration patterns and species interactions. There are five main modes of seed dispersal: gravity, wind, ballistic, water, and animals.
The stability of mutualisms can be undermined by cheating. Cheating is a behavior whereby organisms receive a benefit at the cost of other organisms. Cheating is common in many mutualistic and altruistic relationships (Ferriere et al., 2002). A cheater is an individual who does not cooperate (or cooperates less than their fair share) but can potentially gain the benefit from others cooperating (West et al., 2006). Cheaters are also those who selfishly use common resources to maximize their individual fitness at the expense of a group (McLean & Gudelj, 2006). Simple two-player game theory and the associated evolutionary theory indicate that cheating can be evolutionarily stable, but there are mechanisms to regulate it (Foster & Kokko, 2006).
The mutualism between legume plants and rhizobia (nitrogen-fixing soil bacteria) is an example where this regulation occurs. Rhizobia establish symbiotic organs termed root nodules on the roots of their host, and proliferate by extracting nutrients from the host plant. In turn, they supply their host plants with nitrogen resources produced by nitrogen gas fixation. This mutual nutrient exchange should promote the fitness of both organisms and thereby strengthen the symbiotic relationship. This beneficial effect, known as “partner fidelity feedback”, is assumed as a stabilizing factor for the mutualistic relationship (Sachs et al., 2004; Weyl et al., 2010; Friesen & Jones, 2012; Friesen, 2012).
On the other hand, naturally occurring rhizobium strains vary in their nitrogen fixation activity, and ineffective rhizobia that colonize their host plants without undertaking nitrogen fixation in their root nodules are ubiquitous (Gibson et al., 1975; Bottomly & Jenkins, 1983; Moawad, 1998; Burdon et al., 1999). Because the nitrogen fixation reaction consumes much energy (or costs), such parasitic cheaters could use surplus energy for their own growth or for synthesizing storage substances. Consequently, they are likely to proliferate more efficiently than nitrogen-fixing cooperators, posing a risk to the symbiotic interaction. Rhizobia are therefore exposed to two opposite effects that simultaneously promote (by providing benefit) and destabilize (by incurring cost) the mutualistic relationship.

Figure \(\PageIndex{10}\): Nitrogen-fixing nodules in legumes. Image by Terraprima is licensed under CC BY-SA 3.0.
Despite the widespread presence of ineffective rhizobia, the legume–rhizobia symbiosis is evolutionarily stable. Host sanctioning is a potential mechanism that stabilizes the symbiotic interaction, where plants punish more parasitic cheaters by reducing nutrient supply based on their symbiotic performance (Friesen & Jones, 2012; Friesen, 2012; Kiers & Denison, 1998).
Another example is found within the yucca-yucca moth mutualisms. "Cheating" sometimes happens when the yucca moth deposits too many eggs in one plant. In this case, the yucca plant has little to no benefits from this interaction. However, the plant has a unique way of sanctioning this behavior. While the sanctions against cheating often occurs directly to the individual, in this case, the constraint occurs to the individual's offspring. The yucca plant can "abort" the moths by aborting the flowers. Pellmyr and Huth (1994) found that there is selective maturation for flowers that have low egg loads and high number of scars (and therefore a high amount of pollen). In this way, there is selection against the "cheaters" who try to use the yucca plant without providing the benefits of pollination.

Figure \(\PageIndex{11}\): Two yucca moth females in a yucca flower. The female on the right is laying an egg and clearly shows the pollen ball on the underside of her head. The female on the left is actively pollinating with her maxillary tentacles. Photograph by Olle Pellmyr.
Antagonistic Interactions (Predation, Herbivory, and Parasitism)
Predation is a biological interaction where one organism, the predator, kills and eats another organism, its prey. It is one of a family of common feeding behaviors that includes parasitism and micropredation (which usually do not kill the host) and parasitoidism (which always does, eventually). It is distinct from scavenging on dead prey, though many predators also scavenge. Predation and herbivory overlap because seed predators and destructive frugivores kill their “prey”.
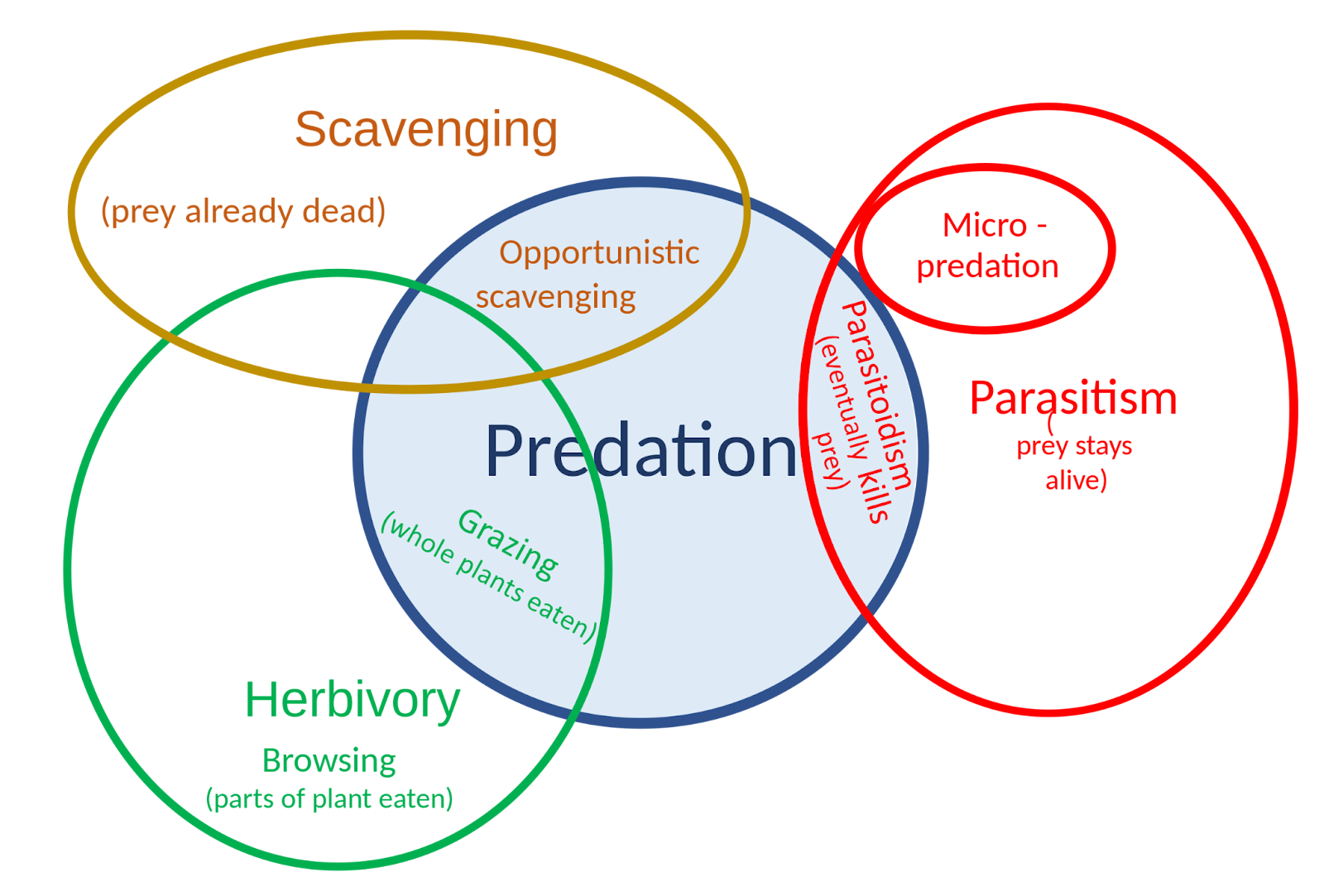
Figure \(\PageIndex{12}\): The relationship of predation to other feeding strategies.
Herbivory is a form of consumption in which an organism principally eats autotrophs such as plants, algae and photosynthesizing bacteria. More generally, organisms that feed on autotrophs in general are known as primary consumers. Herbivory is usually limited to animals that eat plants. Insect herbivory can cause a variety of physical and metabolic alterations in the way the host plant interacts with itself and other surrounding biotic factors (Cannicci et al. 2008; Peschiutta et al. 2018).
Populations of predators and prey in a community are not constant over time: in most cases, they vary in cycles that appear to be related. Population sizes of predators and prey in a community are not constant over time, and they may vary in cycles that appear to be related. The most often cited example of predator-prey dynamics is seen in the cycling of the lynx (predator) and the snowshoe hare (prey), using nearly 100 year-old trapping data from North American forests (Figure \(\PageIndex{13}\)). This cycle of predator and prey lasts approximately years, with the predator population lagging – years behind that of the prey population. As the hare numbers increase, there is more food available for the lynx, allowing the lynx population to increase as well. When the lynx population grows to a threshold level, however, they kill so many hares that hare population begins to decline, followed by a decline in the lynx population because of scarcity of food. When the lynx population is low, the hare population size begins to increase due, at least in part, to low predation pressure, starting the cycle anew.

Predators are adapted and often highly specialized for hunting, with acute senses such as vision, hearing, or smell. Many predatory animals, both vertebrate and invertebrate, have sharp claws or jaws to grip, kill, and cut up their prey. Other adaptations include stealth and aggressive mimicry that improve hunting efficiency. For example, raptors, such as owls and hawks, have hooked beaks for tearing flesh and talons for grabbing prey. Mammal predators often have sharp teeth and claws. Some predators can run quickly to chase prey, and others "sit and wait", lunging forward when prey pass them. Some predators, like rattlesnakes and tarantulas, subdue their prey by injecting them with venom. Predators often have large eyes located forward (like a wolf) rather than eyes spaced far apart on the sides of the head (like a sheep). Forward-facing eyes allow for depth perception, which is key to tracking prey. In contrast, peripheral vision is expanded when eyes are located to the sides, and this helps prey identify threats.
Species have evolved numerous mechanisms to escape predation and herbivory. These defenses may be mechanical, chemical, physical, or behavioral. Some species use their body shape and coloration as camouflage to avoid being detected by predators, an example of a physical defense. The tropical walking stick is an insect with the coloration and body shape of a twig, which makes it very hard to see when it is stationary against a background of real twigs (figure \(\PageIndex{14}\)-a). In another example, the chameleon can change its color to match its surroundings (figure \(\PageIndex{14}\)-b).


Figure \(\PageIndex{14}\): The tropical walking stick and (b) the chameleon use body shape and/or coloration to prevent detection by predators (credit a: modification of work by Linda Tanner; credit b: modification of work by Frank Vassen).
Some prey have armor (such as a turtle shell or the bony plates protecting armadillos), a mechanical defense that reduces predation by discouraging physical contact and some plants have thorns to discourage herbivory (Figure \(\PageIndex{15}\)). Many animals produce or obtain chemical defenses from plants and store them to prevent predation.

Running from predators, hiding, and playing dead are examples of behavioral defenses. Some prey also exhibit behaviors that threaten predators. For example, snapping turtles stretch their legs to appear larger and snap aggressively at predators.
Parasitism is a relationship between species, where one organism, the parasite, lives on or in another organism, the host, causing it some harm, and is adapted structurally to this way of life (Poulin 2007). The parasite either feeds on the host, or, in the case of intestinal parasites, consumes some of its food (Martin and Schwab 2013). Like predation, parasitism is a type of consumer-resource interaction (Getz, 2011), but unlike predators, parasites, with the exception of parasitoids, are typically much smaller than their hosts, do not kill them, and often live in or on their hosts for an extended period. Parasites of animals are highly specialized, and reproduce at a faster rate than their hosts. Classic examples include interactions between vertebrate hosts and tapeworms, flukes, the malaria-causing Plasmodium species, and fleas. Not all parasites kill their hosts, but some do. Parasites increase their own fitness by exploiting hosts for resources necessary for their survival, in particular by feeding on them and by using intermediate (secondary) hosts to assist in their transmission from one definitive (primary) host to another.
The reproductive cycles of parasites are often very complex, sometimes requiring more than one host species. A tapeworm is a parasite that causes disease in humans when contaminated, undercooked meat such as pork, fish, or beef is consumed (Figure \(\PageIndex{16}\)). The tapeworm can live inside the intestine of the host for several years, benefiting from the food the host is bringing into its gut by eating, and may grow to be over 50ft long by adding segments. The parasite moves from species to species in a cycle, making two hosts necessary to complete its life cycle. Another common parasite is Plasmodium falciparum, the protozoan cause of malaria, a significant disease in many parts of the world. Living in human liver and red blood cells, the organism reproduces asexually in the gut of blood-feeding mosquitoes to complete its life cycle. Thus malaria is spread from human to human by mosquitoes, one of many arthropod-borne infectious diseases.

Disease – is a pathological relationship in which the health of plants or animals suffers from an infestation of another species, usually a microbe. Virulent diseases can cause enormous changes in the composition of ecological communities. In the early s, the American chestnut (Castanea dentata) was afflicted by chestnut blight (Endothia parasitica), an introduced fungal pathogen. Because chestnuts have little immunity to this disease, the species was virtually eliminated from the forests of eastern North America by the s. This change released other tree species from competition with the previously dominant chestnut, and they quickly filled gaps in the canopy created by its demise.
A parasitoid, such as an ichneumon wasp, lays its eggs in or on its host; the eggs hatch into larvae, which eat the host, and it inevitably dies. Zoologists generally call this a form of parasitism, though conventionally parasites are thought not to kill their hosts. A predator can be defined to differ from a parasitoid in that it has many prey, captured over its lifetime, where a parasitoid's larva has just one, or at least has its food supply provisioned for it on just one occasion (Lafferty & Kuris, 2002; Gurr & Wratten, 2012).

Figure \(\PageIndex{17}\): Spider wasps paralyze and eventually kill their hosts, but are considered parasitoids, not predators.
Other Types of Interactions
Amensalism
Amensalism (a term introduced by Haskell; Toepfer) is an interaction where an organism inflicts harm to another organism without any costs or benefits received by itself (Willey et al. 2013). Amensalism describes the adverse effect that one organism has on another organism. A classic example of amensalism is where sheep or cattle trample grass. Whilst the presence of the grass causes negligible detrimental effects to the animal's hoof, the grass suffers from being crushed.
Amensalism is often used to describe strongly asymmetrical competitive interactions, such as has been observed between the Spanish ibex and weevils of the genus Timarcha which feed upon the same type of shrub. Whilst the presence of the weevil has almost no influence on food availability, the presence of ibex has an enormous detrimental effect on weevil numbers, as they consume significant quantities of plant matter and incidentally ingest the weevils upon it (Gómez and González-Megías 2002).
Commensalism
Commensalism benefits one organism while the other organism neither benefits nor is harmed. A good example is a remora living with a manatee. Remoras feed on the manatee's feces, and therefore benefits from this interaction. The manatee is not affected by this interaction, as the remora does not deplete the manatee's resources (Williams and Williams 2003). Birds nesting in trees provide an example of a commensal relationship (Figure \(\PageIndex{18}\)). The tree is not harmed by the presence of the nest among its branches. The nests are light and produce little strain on the structural integrity of the branch, and most of the leaves, which the tree uses to get energy by photosynthesis, are above the nest so they are unaffected. The bird, on the other hand, benefits greatly. If the bird had to nest in the open, its eggs and young would be vulnerable to predators. Many potential commensal relationships are difficult to identify because it is difficult to prove that one partner does not derive some benefit from the presence of the other.

Figure \(\PageIndex{18}\): The southern masked-weaver is starting to make a nest in a tree in Zambezi Valley, Zambia. This is an example of a commensal relationship, in which one species (the bird) benefits, while the other (the tree) neither benefits nor is harmed. "African Masked Weaver" by Hanay is licensed under CC BY 3.0.
Community Structure
Communities are complex entities that can be characterized by their structure (the types and numbers of species present) and dynamics (how communities change over time). Understanding community structure and dynamics enables community ecologists to manage ecosystems more effectively. There are several terms for types of communities within communities including indicator species, endemic species, foundation species, keystone species, and invasive species.
Indicator species are environmentally sensitive species that can be used as an early warning system of damage to the ecosystem.
Endemic species are species found only in one geographic area. For example, the blue jay is endemic to North America, while the Barton Springs salamander is endemic to the mouth of one spring in Austin, Texas. Endemics with highly restricted distributions, like the Barton Springs salamander, are particularly vulnerable to extinction. If a population of a widespread species declines in one region, individuals from another region may be able to recolonize the first location, but this is not possible for endemic species. Endemic species are particularly common in isolated regions, such as mountaintops or islands. Endemism is especially likely on islands that are large and far from the mainland.
Foundation Species
Foundation species are considered the “base” or “bedrock” of a community, having the greatest influence on its overall structure. They are usually the primary producers: organisms that bring most of the energy into the community. Kelp, brown algae, is a foundation species, forming the basis of the kelp forests off the coast of California. Foundation species may physically modify the environment to produce and maintain habitats that benefit the other organisms that use them. An example is the photosynthetic corals of the coral reef. Corals themselves are not photosynthetic, but harbor symbionts within their body tissues (dinoflagellates called zooxanthellae) that perform photosynthesis; this is another example of a mutualism. The exoskeletons of living and dead coral make up most of the reef structure, which protects many other species from waves and ocean currents.

Figure \(\PageIndex{19}\): Coral is the foundation species of coral reef ecosystems (credit: Jim E. Maragos, USFWS).

Figure \(\PageIndex{20}\): Foundational species increase food web complexity by facilitating species higher in the food chain. (A) Seven ecosystems with foundation species were sampled: coastal (seagrass, blue mussel, cordgrass), freshwater (watermilfoil, water-starwort) and terrestrial (Spanish moss, marram grass). (B) Food webs were constructed for both bare and foundation species-dominated replicate areas. (C) From each foundation species structured-food web, nodes (species) were randomly removed until the species number matched the species number of the bare food webs.
Although we often focus on trophic (food-related) interactions among species, there is growing evidence that non-trophic interactions can indirectly affect food web topology and trophic dynamics by affecting the species in the network and the strength of trophic links (Sanders et al. 2014; Kefi et al. 2015; van der Zee et al. 2016). Some examples of non-trophic interactions are habitat modification and competition for space.
Foundation species are spatially dominant habitat-structuring organisms (Dayton 1972; Governar 2010; Angelini et al. 2011). Although foundation species are part of the food web like any other species (e.g. as prey or predator), numerous studies have shown that they strongly facilitate the associated community by creating new habitat and alleviating physical stress (Bertness et al. 1999; Jones et al. 2010; Reid and Lortie 2012; Angelini and Silliman 2014; Kefi et al. 2015; van der Zee EM et al. 2015; van der Zee et al. 2016; Filazzola et al. 2017). This form of non-trophic facilitation by foundation species has been found to occur across a wide range of ecosystems and environmental conditions (Bertness and Callaway 1994; Bruno et al. 2003). In harsh coastal zones, corals, kelps, mussels, oysters, seagrasses, mangroves, and salt marsh plants facilitate organisms by attenuating currents and waves, providing aboveground structure for shelter and attachment, concentrating nutrients, and/or reducing desiccation stress during low tide exposure (Bertness and Callaway 1992; Angelini et al. 2011). In more benign systems, foundation species such as the trees in a forest, shrubs and grasses in savannahs, and macrophytes in freshwater systems, have also been found to play a major habitat-structuring role (Jeppesen et al. 1992; Bertness and Callaway 1994; Bruno et al. 2003; Ellison et al. 2005). Ultimately, all foundation species increase habitat complexity and availability, thereby partitioning and enhancing the niche space available to other species (Bruno et al. 2003; Bulleri et al. 2016; Borst et al. 2018).
Borst et al. (2018) tested the general hypothesis that foundation species modify food webs by enhancing their size as indicated by species number, and their complexity as indicated by link density, via facilitation of species, regardless of ecosystem type (Figure \(\PageIndex{20}\)). Additionally, Borst et al. 2018 examined whether any change in food web properties caused by foundation species occurs via random facilitation of species throughout the entire food web or via targeted facilitation of specific species that belong to certain trophic levels or functional groups. They found that species at the base of the food web are less strongly facilitated, and carnivores are more strongly facilitated, in foundation species' food webs than predicted based on random facilitation, resulting in a higher mean trophic level and a longer average chain length. This indicates foundation species strongly enhance food web complexity through non-trophic facilitation of species across the entire trophic network (Borst et al. 2018).
Keystone Species
A keystone species is one whose presence is key to maintaining biodiversity within an ecosystem and to upholding an ecological community’s structure. The intertidal sea star, Pisaster ochraceus, of the northwestern United States is a keystone species (Figure \(\PageIndex{18}\)). Studies have shown that when this organism is removed from communities, populations of their natural prey (mussels) increase, completely altering the species composition and reducing biodiversity. Another keystone species is the banded tetra, a fish in tropical streams, which supplies nearly all of the phosphorus, a necessary inorganic nutrient, to the rest of the community. If these fish were to become extinct, the community would be greatly affected. Keystone species have a greater effect on all other species in the ecosystem, but they don't make up the majority of the biomass. They usually act on communities by changing the food web.

Figure \(\PageIndex{21}\): The Pisaster ochraceus sea star is a keystone species (credit: Jerry Kirkhart).
Invasive species are non-native organisms that, when introduced to an area out of their native range, threaten the ecosystem balance of that habitat. Many such species exist in the United States, as shown in Figure \(\PageIndex{22}\) below. Whether enjoying a forest hike, taking a summer boat trip, or simply walking down an urban street, you have likely encountered an invasive species.

Figure \(\PageIndex{22}\): In the United States, invasive species like (a) purple loosestrife (Lythrum salicaria) and the (b) zebra mussel (Dreissena polymorpha) threaten certain aquatic ecosystems. Some forests are threatened by the spread of (c) common buckthorn (Rhamnus cathartica), (d) garlic mustard (Alliaria petiolata), and (e) the emerald ash borer (Agrilus planipennis). The (f) European starling (Sturnus vulgaris) may compete with native bird species for nest holes. (credit a: modification of work by Liz West; credit b: modification of work by M. McCormick, NOAA; credit c: modification of work by E. Dronkert; credit d: modification of work by Dan Davison; credit e: modification of work by USDA; credit f: modification of work by Don DeBold)
One of the many recent proliferations of an invasive species concerns the growth of Asian carp populations. Asian carp were introduced to the United States in the 1970s by fisheries and sewage treatment facilities that used the fish’s excellent filter feeding capabilities to clean their ponds of excess plankton. Some of the fish escaped, however, and by the 1980s they had colonized many waterways of the Mississippi River basin, including the Illinois and Missouri Rivers.
Voracious eaters and rapid reproducers, Asian carp may outcompete native species for food, potentially leading to their extinction. For example, black carp are voracious eaters of native mussels and snails, limiting this food source for native fish species. Silver carp eat plankton that native mussels and snails feed on, reducing this food source by a different alteration of the food web. In some areas of the Mississippi River, Asian carp species have become the most predominant, effectively outcompeting native fishes for habitat. In some parts of the Illinois River, Asian carp constitute 95 percent of the community's biomass. Although edible, the fish is bony and not a desired food in the United States. Moreover, their presence threatens the native fish and fisheries of the Great Lakes, which are important to local economies and recreational anglers. Asian carp have even injured humans. The fish, frightened by the sound of approaching motorboats, thrust themselves into the air, often landing in the boat or directly hitting the boaters.
The Great Lakes and their prized salmon and lake trout fisheries are also being threatened by these invasive fish. Asian carp have already colonized rivers and canals that lead into Lake Michigan. One infested waterway of particular importance is the Chicago Sanitary and Ship Channel, the major supply waterway linking the Great Lakes to the Mississippi River. To prevent the Asian carp from leaving the canal, a series of electric barriers have been successfully used to discourage their migration; however, the threat is significant enough that several states and Canada have sued to have the Chicago channel permanently cut off from Lake Michigan. Local and national politicians have weighed in on how to solve the problem, but no one knows whether the Asian carp will ultimately be considered a nuisance, like other invasive species such as the water hyacinth and zebra mussel, or whether it will be the destroyer of the largest freshwater fishery of the world.
The issues associated with Asian carp show how population and community ecology, fisheries management, and politics intersect on issues of vital importance to the human food supply and economy. Socio-political issues like this make extensive use of the sciences of population ecology (the study of members of a particular species occupying a particular area known as a habitat) and community ecology (the study of the interaction of all species within a habitat).
Biodiversity, Species Richness, and Relative Species Abundance
Biodiversity describes a community’s biological complexity: it is measured by the number of different species (species richness) in a particular area and their relative abundance (species evenness). The area in question could be a habitat, a biome, or the entire biosphere. Species richness is the term that is used to describe the number of species living in a habitat or biome. Species richness varies across the globe Figure \(\PageIndex{23}\) . One factor in determining species richness is latitude, with the greatest species richness occurring in ecosystems near the equator, which often have warmer temperatures, large amounts of rainfall, and low seasonality. The lowest species richness occurs near the poles, which are much colder, drier, and thus less conducive to life in Geologic time (time since glaciations). The predictability of climate or productivity is also an important factor. Other factors influence species richness as well. For example, the study of island biogeography attempts to explain the relatively high species richness found in certain isolated island chains, including the Galápagos Islands that inspired the young Darwin. Species evenness or Relative species abundance is the number of individuals in a species relative to the total number of individuals in all species within a habitat, ecosystem, or biome. Foundation species often have the highest relative abundance of species.


One factor in determining species richness is latitude, with the greatest species richness occurring in ecosystems near the equator. We call these decreases in biodiversity from equatorial to polar regions the Latitudinal Biodiversity Gradient. The primary cause for the Latitudinal Biodiversity Gradient is climate. Tropical regions which often have warmer temperatures, large amounts of rainfall, and low seasonality (variability between seasons), resulting in species with highly restricted environmental tolerances and limited dispersal ability across environmental barriers. Because of the climate, the tropics also have higher productivity which may lead to larger viable population sizes of specialist species and higher plant diversity. In contrast, the poles, are much colder, drier, and have stronger seasonal climate variability, so are less conducive to life and have lower productivity. Two other possible themes to explain higher tropical rates of net diversification are geographical, pertaining to the greater extent of the tropics being able to support more species compared to other regions, and historical, where the tropics have been less perturbed in the past by climatic events over geologic time (i.e. Pleistocene glaciations). The idea is that greater age provides more time for speciation. The complexity of tropical ecosystems may promote speciation by increasing the heterogeneity, or number of ecological niches, in the tropics relative to higher latitudes. Other factors influence species richness as well.
Two locations with the same richness do not necessarily have the same species evenness. For example, both communities in figure \(\PageIndex{24}\) have three different trees species and thus a species richness of three. However, there is a dominant species (represented by six individuals) in community #. In community #, there are three of individuals of each species. Therefore, community # has a greater species evenness and greater species diversity overall.


Figure \(\PageIndex{24}\): Two hypothetical tree communities have the same species richness, but community #2 (bottom) has a greater species evenness. Both communities have nine trees and three tree species. In community #1, one species is dominant, represented by six individuals. There are two individuals of a conifer species, and only one individual of the final species. In community #2, there are three individuals from each species. Images compiled by Melissa Ha from Alone tree George Hodan, Old Tree Silhouette, and Tree (all public domain).
Species richness can be described in at different scales using alpha, beta, and gamma diversity. Alpha diversity (or species richness), the most commonly referenced measure of species diversity, refers to the total number of species found in a particular biological community, such as a lake or a forest. Bwindi Forest in Uganda, with an estimated 350 bird species, has one of the highest alpha diversities of all African ecosystems. Gamma diversity describes the total number of species that occur across an entire region, such as a mountain range or continent, that includes many ecosystems. The Albertine Rift, which includes Bwindi Forest, has more than 1,074 species of birds, a very high gamma diversity for such a small region. Beta diversity connects alpha and gamma diversity. It describes the rate at which species composition changes across a region. For example, if every wetland in a region was inhabited by a similar suite of plant species, then the region would have low beta diversity; in contrast, if several wetlands in a region had plants communities that were distinct and had little overlap with one another, the region would have high beta diversity. Beta diversity is calculated as gamma diversity divided by alpha diversity. The beta diversity for forest birds of the Albertine Rift is about 3.0, if each ecosystem in the area has about the same number of species as Bwindi Forest.

Figure \(\PageIndex{25}\): Biodiversity indices for nine mountain peaks across three ecoregions. Each symbol represents a different species; some species have populations on only one peak, while others are found on two or more peaks. The variation in species richness on each peak results in different alpha, gamma, and beta diversity values for each ecoregion. This variation has implications for how we divide limited resources to maximize protection. If only one ecoregion can be protected, ecoregion 3 may be a good choice because it has high gamma (total) diversity. However, if only one peak can be protected, should a peak in ecoregion 1 (with many widespread species) or ecoregion 3 (with several unique, range-restricted species) be protected? After Primack, 2012, CC BY 4.0
In 1988, British environmentalist Norman Myers developed a conservation concept to identify areas rich in species and at significant risk for species loss: biodiversity hotspots. Biodiversity hotspots are geographical areas that contain high numbers of endemic species. The purpose of the concept was to identify important locations on the planet for conservation efforts, a kind of conservation triage. By protecting hotspots, governments are able to protect a larger number of species. The original criteria for a hotspot included the presence of 1500 or more endemic plant species and 70 percent of the area disturbed by human activity. There are now 34 biodiversity hotspots (Figure \(\PageIndex{26}\)) containing large numbers of endemic species, which include half of Earth’s endemic plants.

In addition to species diversity, biologists have also identified alternate measures of biodiversity, some of which are important for planning how to preserve biodiversity. Genetic diversity (or variation) is the number of genetic traits within a species and is the raw material for adaptation in a species. A species’ future potential for adaptation depends on the genetic diversity held in the genomes of the individuals in populations that make up the species. The more genetic variations there are within a species, the more likely some members will be able to survive after changes in the environment (leading to adaptation of the species). Most genes code for proteins, which in turn carry out the metabolic processes that keep organisms alive and reproducing. Genetic diversity can be measured as chemical diversity in that different species produce a variety of chemicals in their cells, both the proteins as well as the products and byproducts of metabolism. This chemical diversity is important for humans because of the potential uses for these chemicals, such as medications. For example, the drug eptifibatide is derived from rattlesnake venom and is used to prevent heart attacks in individuals with certain heart conditions. At present, it is far cheaper to discover compounds made by an organism than to imagine them and then synthesize them in a laboratory.
Humans have generated diversity in domestic animals, plants, and fungi through selective breeding. But even this diversity is suffering losses because of market forces and increasing globalism in human agriculture and migration, especially in heavily populated regions such as China, India, and Japan. For example, international seed companies produce only a very few varieties of a given crop and provide incentives around the world for farmers to buy these few varieties while abandoning their traditional varieties, which are far more diverse. The human population depends on crop diversity directly as a stable food source and its decline is troubling to biologists and agricultural scientists.
It is also useful to define ecosystem diversity, meaning the number of different ecosystems on the planet or in a given geographic area. Whole ecosystems can disappear even if some of the species might survive by adapting to other ecosystems. The loss of an ecosystem means the loss of interactions between species, the loss of unique features of coadaptation, and the loss of biological productivity that an ecosystem is able to create. An example of a largely extinct ecosystem in North America is the prairie ecosystem. Prairies once spanned central North America from the boreal forest in northern Canada down into Mexico. They are now all but gone, replaced by crop fields, pasture lands, and suburban sprawl. Many of the species survive, but the hugely productive ecosystem that was responsible for creating the most productive agricultural soils is now gone. As a consequence, soils are disappearing or must be maintained at greater expense. The decline in soil productivity occurs because the interactions in the original ecosystem have been lost; this was a far more important loss to humans than the relatively few species that were driven extinct when the prairie ecosystem was destroyed.
Spatial variation of the environment reflects the fact that conditions are always changing from place to place, and sometimes extremely so. These spatial variations influence the character of ecological communities, in ways that may be gradual or more rapid:
- Gradual changes environmental conditions are associated with varying altitude on a mountain, differences of climate over large distances across continents, and other relatively continuous gradients. This type of spatial change is reflected in gradual variations of communities because individual species have different but overlapping tolerances and requirements of environmental conditions. These biological differences result in overlaps of the distributions of species, which can make it difficult for ecologists to determine the locations of boundaries (ecotones) between types of communities.
- Rapid changes in environmental conditions occur at sharp boundaries between different kinds of soil or bedrock, at interfaces between aquatic and terrestrial habitats, and in places affected by disturbance. The latter influence can occur, for instance, between a burned and unburned tract of forest, or between an ecological reserve and its surrounding area, which may be affected by agriculture or forestry. Relatively discrete changes in environmental conditions favour large differences in community types, with distinct boundaries between them.
Community Dynamics
Community dynamics are the changes in community structure and composition over time. Sometimes these changes are induced by environmental disturbances such as volcanoes, earthquakes, storms, fires, and climate change. Communities with a stable structure are said to be at equilibrium. Following a disturbance, the community may or may not return to the equilibrium state.
Disturbance is an event of destruction of some part of a community, an occurrence that is followed by a sometimes pronged period of ecological recovery called succession. All communities are dynamic, changing over time in their species composition and functional attributes (such as productivity, decomposition, and nutrient cycling). However, the rate of change depends on the stability of environmental conditions, which is greatest in communities that are close to the end-point of a succession. In contrast, the most dynamic communities are associated with the younger stages of succession. Disturbances can occur on two spatial scales.
- Stand-replacing disturbances are caused by wildfire, a disease epidemic, clear-cutting, and other cataclysmic events (Figure \(\PageIndex{27}\)). This kind of disturbance is extensive and results in the immediate replacement of a community with a different one, followed by a period of successional recovery. Over time, succession may regenerate a community similar to what existed before the disruption, or a different one may result. The younger stages of a sere (successional sequence) are especially dynamic in terms of community change.
- Microdisturbances are local disruptions that affect small areas within an otherwise intact community. A microdisturbance may, for instance, be associated with the death of an individual large tree, which results in a gap in the canopy, below which community change is relatively dynamic as species compete to take advantage of the additional sunlight. Similarly, the death of an individual coral head represents a microdisturbance within a tropical reef community. Although ecological changes are dynamic within a gap created by a recent microdisturbance, at the stand level the community is relatively stable. Gap-phase community dynamics occur in all ecosystems but are especially important during later stages of succession, such as in older-growth forests.


Figure \(\PageIndex{27}\): Ecosystems are occasionally subjected to catastrophic disturbances, such as these forest fires in in the boreal forest of northern Quebec. The individual fires are marked with a red dot, and their smoke plumes are blowing to the south. The large white mass at the bottom right is cloud cover. Source: NASA photo ID ; https://www.dvidshub.net/image//fires-quebec-canada-send-smoke-us-natural-hazards#.VOS_vXUtHIU
Succession describes the sequential appearance and disappearance of species in a community over time. In primary succession, newly exposed or newly formed land is colonized by living things; in secondary succession, part of an ecosystem is disturbed and remnants of the previous community remain.

Figure \(\PageIndex{28}\): During primary succession in lava on Maui, Hawaii, succulent plants are the pioneer species (credit: Forest and Kim Starr).
Primary succession occurs following the eruption of volcanoes, such as those on the Big Island of Hawaii. As lava flows into the ocean, new land is continually being formed. On the Big Island, approximately 32 acres of land is added each year. First, weathering and other natural forces break down the substrate enough for the establishment of certain hearty plants and lichens with few soil requirements, known as pioneer species (Figure \(\PageIndex{28}\)). These species help to further break down the mineral rich lava into soil where other, less hardy species will grow and eventually replace the pioneer species. In addition, as these early species grow and die, they add to an ever-growing layer of decomposing organic material and contribute to soil formation. Over time the area will reach an equilibrium state, with a set of organisms quite different from the pioneer species.

Figure \(\PageIndex{29}\): Trajectory of forest secondary succession following an intense disturbance, resulting in nearly no original biomass, to a mature forest state. Biodiversity, biomass, and soil depth all increase with time, and community composition changes. Image by Lucas Martin Frey is licensed under CC BY 3.0.
A classic example of secondary succession occurs in oak and hickory forests cleared by wildfire (Figure \(\PageIndex{30}\)). Wildfires will burn most vegetation and kill those animals unable to flee the area. Their nutrients, however, are returned to the ground in the form of ash. Thus, even when areas are devoid of life due to severe fires, the area will soon be ready for new life to take hold. Before the fire, the vegetation was dominated by tall trees with access to the major plant energy resource: sunlight. Their height gave them access to sunlight while also shading the ground and other low-lying species. After the fire, though, these trees are no longer dominant. Thus, the first plants to grow back are usually annual plants followed within a few years by quickly growing and spreading grasses and other pioneer species. Due to, at least in part, changes in the environment brought on by the growth of the grasses and other species, over many years, shrubs will emerge along with small pine, oak, and hickory trees. These organisms are called intermediate species. Eventually, over 150 years, the forest will reach its equilibrium point where species composition is no longer changing and resembles the community before the fire.
Unlike in primary succession, the species that dominate secondary succession, are usually present from the start of the process, often in the soil seed bank. In some systems the successional pathways are fairly consistent, and thus, are easy to predict. Ecological succession was formerly seen as having a stable end-stage called the climax, sometimes referred to as the 'potential vegetation' of a site, and shaped primarily by the local climate. This idea has been largely abandoned by modern ecologists in favor of non-equilibrium ideas of ecosystems dynamics. In others, there are many possible pathways, potentially leading to alternative stable states. For example, nitrogen-fixing legumes alter successional trajectories (Chapin et al., 2002). Most natural ecosystems experience disturbance at a rate that makes a "climax" community unattainable. Climate change often occurs at a rate and frequency sufficient to prevent arrival at a climax state. Additions to available species pools through range expansions and introductions can also continually reshape communities.

References
Alexander, R.R., Pond, F.W., & Rodgers, J.E. (n.d.). Yucca (L.). Forest Service Handbooks. https://www.fs.fed.us/rm/pubs_other/wo_AgricHandbook727/wo_AgricHandbook727_1175_1177.pdf
Bottomley. P, & Jenkins, M. (1983). Some characteristic of Rhizobium meliloti isolates from alfalfa fields in Oregon. Soil Sci. Soc. Am., J 47, pp. 1153–1157.
Burdon, J., Gibson, A., Searle, S., Woods, M., & Brockwell, J. (1999). Variation in the effectiveness of symbiotic associations between native rhizobia and temperate Australian Acacia: Within-species interactions. J. Appl. Ecol., 36, pp. 398–408.
Ferriere, R., Bronstein, J.L., Rinaldi, S., Law, R., & Gauduchon, M. (2002). Cheating and the evolutionary stability of mutualisms. Proc. R. Soc. Lond., 269(1493), pp. 773–780. doi:10.1098/rspb.2001.1900. PMC 1690960. PMID 11958708.
Foster, K.R., & Kokko, H. (2006). Cheating can stabilize cooperation in mutualisms. Proceedings of the Royal Society B: Biological Sciences, 273(1598), pp. 2233-2239. doi:10.1098/rspb.2006.3571. PMC 1635526. PMID 16901844.
Friesen, M.L., & Jones, E.I. (2012). Modelling the evolution of mutualistic symbioses. Methods Mol. Biol., 804, pp. 481–499.
Friesen, M.L. (2012). Widespread fitness alignment in the legume-rhizobium symbiosis. New Phytol., 194, pp. 1096–1111.
Gibson, A., Curnow, B., Bergersen, F., Brockwell, J., & Robinson, A. (1975). Studies of field populations of Rhizobium: Effectiveness of strains of Rhizobium trifolii associated with Trifolium subterraneum L. pastures in South-Eastern Australia. Soil Biol. Biochem., 7, pp. 95–102.
Kiers, E., & Denison, R. (2008). Sanctions, cooperation, and the stability of plant Rhizosphere mutualisms. Annu. Rev. Ecol. Evol. Syst., 39, pp. 215–236.
MacLean, R.C., & Gudelj, I. (2006). Resource competition and social conflict in experimental populations of yeast. Nature, 441(7092), 498-501. Bibcode:2006Natur.441..498M. doi:10.1038/nature04624. PMID 16724064. S2CID 4419943.
Moawad, H., El-Din, S., & Abdel-Aziz, R. (1998). Improvement of biological nitrogen fixation in Egyptian winter legumes through better management of Rhizobium. Plant Soil, 204, pp. 95–106.
Pellmyr, O., & Huth, C.J. (1994). Evolutionary stability of mutualism between yuccas and yucca moths. Nature, 372(6503), pp. 257–260. Bibcode:1994Natur.372..257P. doi:10.1038/372257a0. S2CID 4330563.
Sachs, J., Mueller, U., Wilcox, T., & Bull, J. (2004). The evolution of cooperation. Quart. Rev. Biol., 79, pp. 135–160.
West, S.A., Griffin, A.S., Gardner, A., & Diggle, S.P. (2006). Social evolution theory for microorganisms. Nature Reviews Microbiology, 4(8), pp. 597-607. doi:10.1038/nrmicro1461. PMID 16845430. S2CID 18451640.
Weyl, E.G., Frederickson, M.E., Yu, D.W., & Pierce, N.E. (2010). Economic contract theory tests models of mutualism. Proc. Natl. Acad. Sci. USA., 107, pp. 15712–15716.
Predator & prey: Adaptations. (2012). Royal Saskatchewan Museum. Archived from the original (PDF) on 3 April 2018. Retrieved 19 April 2018.
"Types of Pollination, Pollinators and Terminology". CropsReview.Com. Retrieved 2015-10-20.
Angelini, C., Altieri, A.H., Silliman, B.R., & Bertness, M.D. (2011). Interactions among foundation species and their consequences for community organization, biodiversity, and conservation. Bioscience, 61(10), pp. 782–9.
Angelini, C., & Silliman, B.R. (2014). Secondary foundation species as drivers of trophic and functional diversity: Evidence from a tree epiphyte system. Ecology, 95(1), pp. 185–96. pmid:24649658
Bar-Yam. Predator-prey relationships. New England Complex Systems Institute. Retrieved 7 September 2018.
Begon, M., Harper, J.L., & Townsend, C.R. (1996). Ecology: Individuals, populations, and communities, Third Edition. Blackwell Science Ltd., Cambridge, Massachusetts, USA.
Bengtson, S. (2002). Origins and early evolution of predation. In Kowalewski, M., & Kelley, P.H. (eds.). The fossil record of predation. The Paleontological Society Papers 8. The Paleontological Society. pp. 289–317.
Bertness, M.D., & Callaway, R. (1994). Positive interactions in communities. Trends in Ecology & Evolution, 9(5), pp. 191–3.
Bertness, M.D., Leonard, G.H., Levine, J.M., Schmidt, P.R., & Ingraham, A.O. (1999). Testing the relative contribution of positive and negative interactions in rocky intertidal communities. Ecology, 80(8), pp. 2711–26.
Borst, A.C., Verberk, W.C., Angelini, C., Schotanus, J., Wolters, J.W., Christianen, M.J., van der Zee, E.M., Derksen-Hooijberg, M., & van der Heide, T. (2018). Foundation species enhance food web complexity through non-trophic facilitation. PLOS ONE, 13(8): e0199152. doi:10.1371/journal.pone.0199152. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
Bruno, J.F., Stachowicz, J.J., & Bertness, M.D. (2003). Inclusion of facilitation into ecological theory. Trends in Ecology & Evolution, 18(3), pp. 119–25.
Bulleri, F., Bruno, J.F., Silliman, B.R., Stachowicz, J.J. (2016). Facilitation and the niche: Implications for coexistence, range shifts and ecosystem functioning. Functional Ecology, 30(1), pp. 70–8.
Cannicci, S., Burrows, D., Fratini, S., Smith, T.J., Offenberg, J., & Dahdouh-Guebas, D. (2008). Faunal impact on vegetation structure and ecosystem function in mangrove forests: A review. Mangrove Ecology – Applications in Forestry and Costal Zone Management. Aquatic Botany, 89(2), pp. 186–200.
Dayton, P.K. (1972). Toward an understanding of community resilience and the potential effects of enrichments to the benthos at McMurdo Sound, Antarctica. In Parker, B. (Eds.), Proceedings of the colloquium on conservation problems in Antarctica. Lawrence, Kansas: Allen Press.
Ehrlich, P.R., & Raven, P.H. (1964). Butterflies and plants: A study in coevolution. Evolution 18(4), pp. 586–608. doi:10.2307/2406212. JSTOR 2406212.
Ellison, A.M., Bank, M.S., Clinton, B.D., Colburn, E.A., Elliott, K., Ford, C.R., et al. (2005). Loss of foundation species: Consequences for the structure and dynamics of forested ecosystems. Frontiers in Ecology and the Environment, 3(9), pp. 479–86.
Filazzola, A., Westphal, M., Powers, M., Liczner, A.R., Woollett, D.A., Johnson, B., et al. (2017). Non-trophic interactions in deserts: Facilitation, interference, and an endangered lizard species. Basic and Applied Ecology, 20, pp. 51–61.
Gómez, J.M., & González-Megías, A. (2002). Asymmetrical interactions between ungulates and phytophagous insects: Being different matters. Ecology, 83(1), pp. 203–11. doi:10.1890/0012-9658(2002)083[0203:AIBUAP]2.0.CO;2.
Govenar, B. (2010). Shaping vent and seep communities: Habitat provision and modification by foundation species. In Kiel, S., (Eds.), The vent and seep biota. Topics in Geobiology. 33: Springer Netherlands, pp. 403–32.
Hardin, G. (1960). The competitive exclusion principle. Science, 131(3409), pp. 1292–1297. Bibcode:1960Sci...131.1292H. doi:10.1126/science.131.3409.1292. PMID 14399717.
Jeppesen, E., Sondergaard, M., Sondergaard, M., & Christofferson, K. (1992). The structuring role of submerged macrophytes in lakes. New York: Springer.
Jones, C.G., Gutierrez, J.L., Byers, J.E,. Crooks, J.A., Lambrinos, J.G., & Talley, T.S. (2010). A framework for understanding physical ecosystem engineering by organisms. Oikos, 119(12), pp. 1862–9.
Kefi, S., Berlow, E.L., Wieters, E.A., Joppa, L.N., Wood, S.A., Brose, U., et al. Network structure beyond food webs: Mapping non-trophic and trophic interactions on Chilean rocky shores. Ecology, 96(1), pp. 291–303. pmid:26236914.
Lim, G., & Burns, K.C. (2021). Do fruit reflectance properties affect avian frugivory in New Zealand? New Zealand Journal of Botany, pp. 1–11. doi:10.1080/0028825X.2021.2001664. ISSN 0028-825X. S2CID 244683146.
Lunau, K. (2004). Adaptive radiation and coevolution — pollination biology case studies. Organisms Diversity & Evolution, 4(3), pp. 207–224. doi:10.1016/j.ode.2004.02.002.
Martin, B.D., & Schwab, E. (2013). Current usage of symbiosis and associated terminology. International Journal of Biology, 5(1), pp. 32–45. doi:10.5539/ijb.v5n1p32.
Peschiutta, M.L., Scholz, F.G., Goldstein, G., & Bucci, S.J. (2018). Herbivory alters plant carbon assimilation, patterns of biomass allocation and nitrogen use efficiency. Acta Oecologica, 86, pp. 9–16. doi:10.1016/j.actao.2017.11.007.
Pocheville, A. (2015). The ecological niche: History and recent controversies. In Heams, T., Huneman, P., Lecointre, G., et al. (Eds.). Handbook of evolutionary thinking in the sciences. Dordrecht: Springer. pp. 547–586. ISBN 978-94-017-9014-7.
Pollan, M. (2001). The botany of desire: A plant's-eye view of the world. Bloomsbury. ISBN 978-0-7475-6300-6.
Poulin, R. (2007). Evolutionary ecology of parasites. Princeton University Press, pp. 4–5. ISBN 978-0-691-12085-0.
Reid, A.M., & Lortie, C.J. (2012). Cushion plants are foundation species with positive effects extending to higher trophic levels. Ecosphere, 3(11).
Sahney, S., Benton, M.J., & Ferry, P.A. (2010). Links between global taxonomic diversity, ecological diversity and the expansion of vertebrates on land. Biology Letters, 6(4), pp. 544–547. doi:10.1098/rsbl.2009.1024. PMC 2936204. PMID 20106856.
Sanders, D., Jones, C.G., Thébault, E., Bouma, T.J., Heide Tvd, Belzen Jv, et al. (2014). Integrating ecosystem engineering and food webs. Oikos, 123(5), pp. 513–24.
Toepfer, G. "Amensalism". In: BioConcepts. link.
Van der Zee, E.M., Angelini, C., Govers, L.L., Christianen, M.J.A., Altieri, A.H., van der Reijden, K.J., et al. (2016). How habitat-modifying organisms structure the food web of two coastal ecosystems. Proceedings of the Royal Society B-Biological Sciences, 283(1826). pmid:26962135.
Van der Zee, E.M., Tielens, E., Holthuijsen, S., Donadi, S., Eriksson, B.K., van der Veer, H.W., et al. (2015). Habitat modification drives benthic trophic diversity in an intertidal soft-bottom ecosystem. Journal of Experimental Marine Biology and Ecology, 465, pp. 41–8.
Vermeij, G.J. (1993). Evolution and escalation: An ecological history of life. Princeton University Press. pp. 11 and passim. ISBN 978-0-691-00080-0.
Willey, J.M., Sherwood, L.M., & Woolverton, C.J. (2013). Prescott's Microbiology (9th ed.). pp. 713–38. ISBN 978-0-07-751066-4.
Williams, E., Mignucci, W.L., & Bonde. (2003). Echeneid-sirenian associations, with information on sharksucker diet. Journal of Fish Biology, 5(63), pp. 1176–1183. doi:10.1046/j.1095-8649.2003.00236.x. Retrieved 17 June 2020.
Wootton, J.T., & Emmerson, M. (2005). Measurement of interaction strength in nature. Annual Review of Ecology, Evolution, and Systematics, 36, pp. 419–44. doi:10.1146/annurev.ecolsys.36.091704.175535. JSTOR 30033811.
Angert, A.L., Huxman, T.E., Chesson, P., & Venable, D.L. (2009). Functional tradeoffs determine species coexistence via the storage effect. Proceedings of the National Academy of Sciences, 106(28), pp. 11641–11645. doi:10.1073/pnas.0904512106
Caldwell, J.P., & Vitt, L.J. (1999). Dietary asymmetry in leaf litter frogs and lizards in a transitional northern Amazonian rain forest. Oikos, 84(3), pp. 383–397. doi:10.2307/3546419
Chase, J.M., Abrams, P.A., Grover, J.P., Diehl, S., Chesson, P., Holt, R.D., Richards, S.A., Nisbet, R.M., & Case, T.J. (2002). The interaction between predation and competition: A review and synthesis. Ecology Letters, 5(2), pp. 302–315. doi:10.1046/j.1461-0248.2002.00315.x
Chesson, P., & Warner, R. (1981). Environmental variability promotes coexistence in lottery competitive systems. The American Naturalist, 117(6), pp. 923–943. doi:10.1086/283778
Chesson, P. (2000). Mechanisms of maintenance of species diversity. Annual Review of Ecology and Systematics, 31, pp. 343–366. doi:10.1146/annurev.ecolsys.31.1.343
Chesson, P., & Kuang, J.J. (2008). The interaction between predation and competition. Nature, 456(7219), pp. 235–238. doi:10.1038/nature07248
Grover, J.P. (1997). Resource competition (1st ed.). London: Chapman & Hall. ISBN 978-0412749308
Hardin, G. (1960). The competitive exclusion principle. Science, 131(3409), pp. 1292–1297. doi:10.1126/science.131.3409.1292
Holt, R.D., Grover, J., & Tilman, D. (1994). Simple rules for interspecific dominance in systems with exploitative and apparent competition. The American Naturalist, 144(5), pp. 741–771. doi:10.1086/285705
Hutchinson, G.E. (1961). The paradox of the plankton. The American Naturalist, 95(882), pp. 137–145. doi:10.1086/282171
MacArthur, R.H. (1958). Population ecology of some warblers of northeastern coniferous forests. Ecology, 39(4), pp. 599-619.
Pacala, S.W., & Roughgarden, J. (1985). Population experiments with the Anolis lizards of St. Maarten and St. Eustatius. Ecology, 66(1), pp. 129–141. doi:10.2307/1941313
Sedio, B.E., Ostling, A.M. (2013). How specialised must natural enemies be to facilitate coexistence among plants? Ecology Letters, 16(8), pp. 995–1003. doi:10.1111/ele.12130
Tilman, D. (1980). Resources: A graphical-mechanistic approach to competition and predation. The American Naturalist, 116(3), pp. 362–393. doi:10.1086/283633
Contributors and Attributions
This chapter was written by K. Whittinghill, N. Gownaris, A. Howard, C. Olmstead, and T. Zallek, with text taken from the following CC-BY resources:
- 45.6: Community Ecology by OpenStax, is licensed CC BY
- Biological Interaction by Wikipedia, the free encyclopedia
- Herbivore by Wikipedia, the free encyclopedia
- Ecosystems by CK-: Biology Concepts, is licensed CC BY-NC
- Essentials of Environmental Science by Kamala Doršner is licensed under CC BY
- Ecology: From Individuals to the Biosphere from Environmental Science: A Canadian Perspective by Bill Freedman
- Community Ecology and Importance of Biodiversity from Environmental Biology by Matthew R. Fisher (licensed under CC-BY)
- Preserving Biodiversity and The Biodiversity Crisis by OpenStax, is licensed CC BY by
Connie Rye (East Mississippi Community College), Robert Wise (University of Wisconsin, Oshkosh), Vladimir Jurukovski (Suffolk County Community College), Jean DeSaix (University of North Carolina at Chapel Hill), Jung Choi (Georgia Institute of Technology), Yael Avissar (Rhode Island College) among other contributing authors. Original content by OpenStax (CC BY 4.0; Download for free at http://cnx.org/contents/185cbf87-c72...f21b5eabd@9.87).
- The Biodiverisity Crisis and Prelude to Conservation Biology and Biodiversity from General Biology by OpenStax (licensed under CC-BY)
- Biodiversity and Biomes from Life Sciences Grade by Siyavula (CC-BY)
- Biological Interaction by Wikipedia, the free encyclopedia
- Herbivore by Wikipedia, the free encyclopedia
- Conservation Biology in Sub-Saharan Africa by John W. Wilson and Richard B. Primack
-
Competitive exclusion principle by Wikipedia, the free encyclopedia
- Cheating (biology) by Wikipedia, the free encyclopedia
- Fujita H, Aoki S, Kawaguchi M (2014) Evolutionary Dynamics of Nitrogen Fixation in the Legume–Rhizobia Symbiosis. PLoS ONE 9(4): e93670. doi:10.1371/journal.pone.0093670
- https://bio.libretexts.org/Courses/L...e_and_Dynamics