
16.1: Predation
- Page ID
- 83705
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)

Figure \(\PageIndex{1}\): Polar bears (top photo) are solitary predators, here shown feeding on a beaded seal. Other species, like meat ants (photo below), cooperate to feed and are called social predators. Here, meat ants feed socially on a cicada far larger than themselves.
Predation is a biological interaction where one organism, the predator, kills and eats another organism, its prey. It is one of a family of common feeding behaviors that includes parasitism and micropredation (which usually do not kill the host) and parasitoidism (which always does, eventually). It is distinct from scavenging on dead prey, though many predators also scavenge. Predation and herbivory overlap because seed predators and destructive frugivores kill their “prey”.
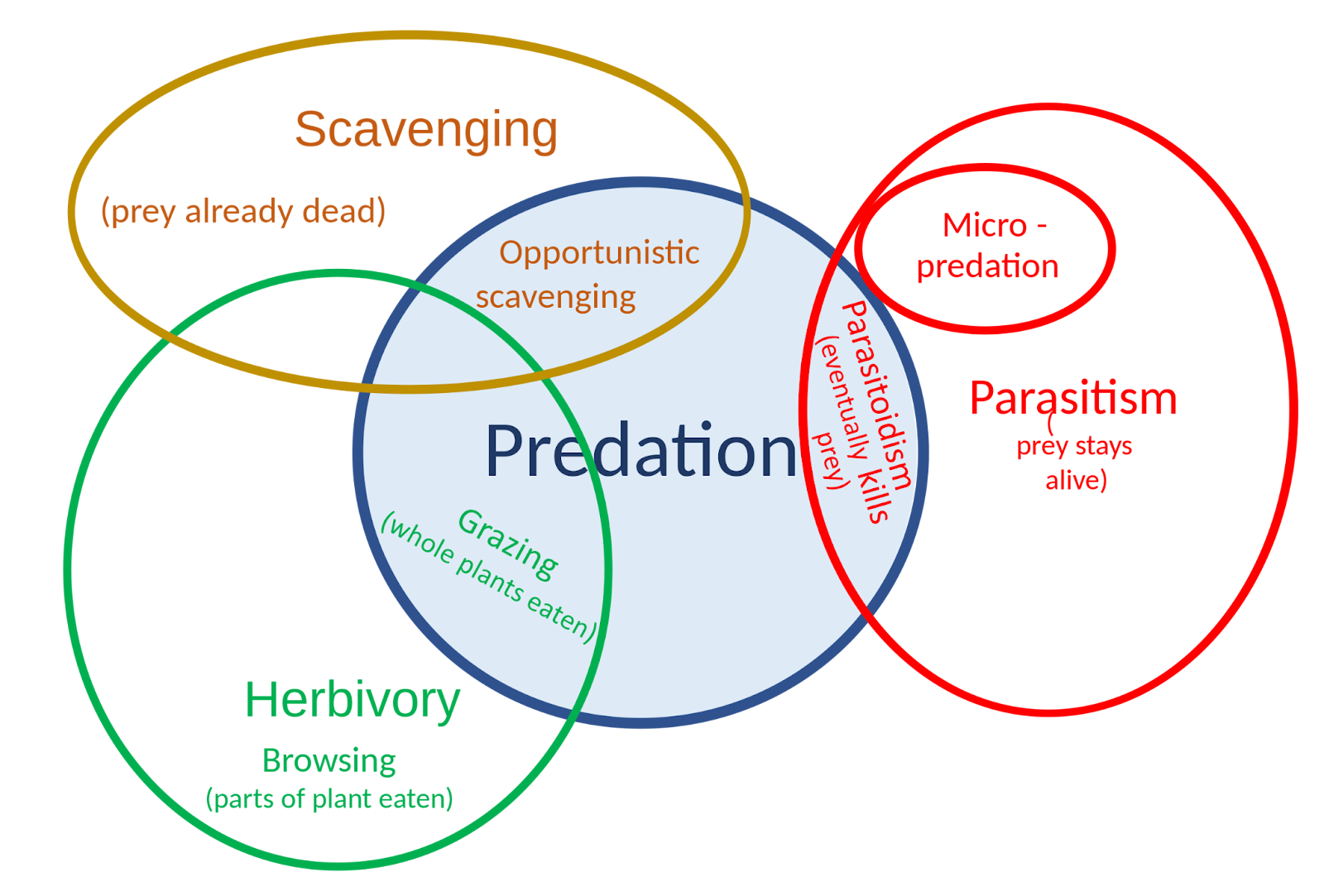
Figure \(\PageIndex{2}\): The relationship of predation to other feeding strategies.
The concept of predation is broad, defined differently in different contexts, and includes a wide variety of feeding methods; and some relationships that result in the prey's death are not generally called predation. A parasitoid, such as an ichneumon wasp, lays its eggs in or on its host; the eggs hatch into larvae, which eat the host, and it inevitably dies. Zoologists generally call this a form of parasitism, though conventionally parasites are thought not to kill their hosts. A predator can be defined to differ from a parasitoid in that it has many prey, captured over its lifetime, where a parasitoid's larva has just one, or at least has its food supply provisioned for it on just one occasion (Lafferty & Kuris, 2002; Gurr & Wratten, 2012).

Figure \(\PageIndex{3}\): Spider wasps paralyze and eventually kill their hosts, but are considered parasitoids, not predators.
There are other difficult and borderline cases. Micropredators are small animals that, like predators, feed entirely on other organisms; they include fleas and mosquitoes that consume blood from living animals, and aphids that consume sap from living plants. However, since they typically do not kill their hosts, they are now often thought of as parasites (Poulin et al., 2011; Poulin & Randhawa, 2015). Animals that graze on phytoplankton or mats of microbes are predators, as they consume and kill their food organisms; but herbivores that browse leaves are not, as their food plants usually survive the assault (Bengston 2002). Scavengers, organisms that only eat organisms found already dead, are not predators, but many predators such as the jackal and the hyena scavenge when the opportunity arises (Kruuk, 1972; Kane et al., 2017). Among invertebrates, social wasps (yellowjackets) are both hunters and scavengers of other insects (Schmidt, 2009).
Predators are adapted and often highly specialized for hunting, with acute senses such as vision, hearing, or smell. Many predatory animals, both vertebrate and invertebrate, have sharp claws or jaws to grip, kill, and cut up their prey. Other adaptations include stealth and aggressive mimicry that improve hunting efficiency.
When prey is detected, the predator assesses whether to attack it. Predators may actively search for or pursue prey (pursuit predation) or sit and wait for prey (ambush predation), often concealed, prior to attack. If the attack is successful, the predator kills the prey, removes any inedible parts like the shell or spines, and eats it.
Predation has a powerful selective effect on prey, and the prey develop antipredator adaptations such as warning coloration, alarm calls and other signals, camouflage, mimicry of well-defended species, and defensive spines and chemicals. Sometimes predator and prey find themselves in an evolutionary arms race, a cycle of adaptations and counter-adaptations.
16.1.1 Foraging

Figure \(\PageIndex{4}\): A basic foraging cycle for a predator, with some variations indicated (Kramer, 2001).
To feed, a predator must search for, pursue and kill its prey. These actions form a foraging cycle (Griffiths, 1980; Wetzel & Liken, 2000). The predator must decide where to look for prey based on its geographical distribution; and once it has located prey, it must assess whether to pursue it or to wait for a better choice. If it chooses pursuit, its physical capabilities determine the mode of pursuit (e.g., ambush or pursuit) (MacArthur, 1984; Pianka, 2011). Having captured the prey, it may also need to expend energy handling it (e.g., killing it, removing any shell or spines, and ingesting it) (Griffiths, 1980; Kramer, 2001).
Search
Predators have a choice of search modes ranging from sit-and-wait to active or widely foraging (Perry, 1999; Kramer, 2011; Bell, 2012; Eastman & Thiel, 2015). The sit-and-wait method is most suitable if the prey are dense and mobile, and the predator has low energy requirements (Bell, 2012). Wide foraging expends more energy, and is used when prey is sedentary or sparsely distributed (Pianka, 2011; Bell, 2012). There is a continuum of search modes with intervals between periods of movement ranging from seconds to months. Sharks, sunfish, insectivorous birds and shrews are almost always moving while web-building spiders, aquatic invertebrates, praying mantises and kestrels rarely move. In between, plovers and other shorebirds, freshwater fish including crappies, and the larvae of coccinellid beetles (ladybirds), alternate between actively searching and scanning the environment (Bell, 2012).

Figure \(\PageIndex{5}\): The black-browed albatross regularly flies hundreds of kilometers across the nearly empty ocean to find patches of food.
Prey distributions are often clumped, and predators respond by looking for patches where prey is dense and then searching within patches (Kramer, 2001). Where food is found in patches, such as rare shoals of fish in a nearly empty ocean, the search stage requires the predator to travel for a substantial time, and to expend a significant amount of energy, to locate each food patch (Bell, 2012). For example, the black-browed albatross regularly makes foraging flights to a range of around 700 kilometers (430 miles), up to a maximum foraging range of 3,000 kilometers (1,860 miles) for breeding birds gathering food for their young (Gremillet et al., 2000). With static prey, some predators can learn suitable patch locations and return to them at intervals to feed (Bell, 2012).
Assessment

Figure \(\PageIndex{6}\): Seven-spot ladybirds select plants of good quality for their aphid prey.
Having found prey, a predator must decide whether to pursue it or keep searching. The decision depends on the costs and benefits involved. A bird foraging for insects spends a lot of time searching but capturing and eating them is quick and easy, so the efficient strategy for the bird is to eat every palatable insect it finds. By contrast, a predator such as a lion or falcon finds its prey easily but capturing it requires a lot of effort. In that case, the predator is more selective (Pianka, 2011).
One of the factors to consider is size. Prey that is too small may not be worth the trouble for the amount of energy it provides. Too large, and it may be too difficult to capture. For example, a mantid captures prey with its forelegs and they are optimized for grabbing prey of a certain size. Mantids are reluctant to attack prey that is far from that size. There is a positive correlation between the size of a predator and its prey (Pianka, 2011).
Capture
To capture prey, predators have a spectrum of pursuit modes that range from overt chase (pursuit predation) to a sudden strike on nearby prey (ambush predation) (Stevens, 2010; Kramer, 2011; Williams et al., 2012)
Ambush


Figure \(\PageIndex{7}\): A western green lizard ambushes its grasshopper prey (top photo) and a trapdoor spider waits in its burrow to ambush its prey (bottom photo).
Ambush or sit-and-wait predators are carnivorous animals that capture prey by stealth or surprise. In animals, ambush predation is characterized by the predator's scanning the environment from a concealed position until a prey is spotted, and then rapidly executing a fixed surprise attack (deVries et al., 2012; Moore & Biewener, 2015). Vertebrate ambush predators include frogs, fish such as the angel shark, the northern pike and the eastern frogfish (Canadian Wildlife Service, 2007; Bray, 2014; Moore & Biewener, 2015; Indiana Division of Fish and Wildlife, 2018). Among the many invertebrate ambush predators are trapdoor spiders and Australian Crab spiders on land and mantis shrimps in the sea (deVries et al., 2012; BBC, 2014; Arizona-Sonora Desert Museum). Ambush predators often construct a burrow in which to hide, improving concealment at the cost of reducing their field of vision. Some ambush predators also use lures to attract prey within striking range (Moore & Biewener, 2015). The capturing movement has to be rapid to trap the prey, given that the attack is not modifiable once launched (Moore & Biewener, 2015).
Pursuit


Figure \(\PageIndex{8}\): Humpback whales (top photo) are lunge feeders, filtering thousands of krill from seawater and swallowing them alive. Dragonflies (bottom photo), like this common clubtail with captured prey, are invertebrate pursuit predators.
In pursuit predation, predators chase fleeing prey. If the prey flees in a straight line, capture depends only on the predator's being faster than the prey (Moore & Biewener, 2015). If the prey maneuvers by turning as it flees, the predator must react in real time to calculate and follow a new intercept path as it closes on the prey (Moore & Biewener, 2015). Many pursuit predators use camouflage to approach the prey as close as possible unobserved (stalking) before starting the pursuit (Moore & Biewener, 2015). Pursuit predators include terrestrial mammals such as humans, African wild dogs, spotted hyenas and wolves; marine predators such as dolphins, orcas and many predatory fishes, such as tuna (Gazda et al., 2005; Tyus, 2011); predatory birds (raptors) such as falcons; and insects such as dragonflies (Combes et al., 2013). Pursuit predators may be social, like the lion and wolf that hunt in groups, or solitary (Lafferty & Kuris, 2002).
An extreme form of pursuit is endurance or persistence hunting, in which the predator tires out the prey by following it over a long distance, sometimes for hours at a time. The method is used by human hunter-gatherers and by canids such as African wild dogs and domestic hounds. The African wild dog is an extreme persistence predator, tiring out individual prey by following them for many miles at relatively low speed (Hubel et al., 2016).
A specialized form of pursuit predation is the lunge feeding of baleen whales. These very large marine predators feed on plankton, especially krill, diving and actively swimming into concentrations of plankton, and then taking a huge gulp of water and filtering it through their feathery baleen plates (Goldbogen et al., 2006; Sanders et al., 2015).
Handling


Figure \(\PageIndex{9}\): Catfish has sharp dorsal and pectoral spines which it holds erect to discourage predators such as herons which swallow prey whole (top photo). Osprey tears its fish prey apart, avoiding dangers such as sharp spines.
Once the predator has captured the prey, it has to handle it: very carefully if the prey is dangerous to eat, such as if it possesses sharp or poisonous spines, as in many prey fish. Some catfish such as the Ictaluridae have spines on the back (dorsal) and belly (pectoral) which lock in the erect position; as the catfish thrashes about when captured, these could pierce the predator's mouth, possibly fatally. Some fish-eating birds like the osprey avoid the danger of spines by tearing up their prey before eating it (Forbes, 1989). Some prey may also be time-consuming to handle, such as clams or other bivalves that are difficult for predators to crush or open.
16.1.2 Predator Adaptations
Physical adaptations
Under the pressure of natural selection, predators have evolved a variety of physical adaptations for detecting, catching, killing, and digesting prey. These include speed, agility, stealth, sharp senses, claws, teeth, filters, and suitable digestive systems (Bar-Yam, 2018).
For detecting prey, predators have well-developed vision, smell, or hearing (Stevens et al., 2010). Predators as diverse as owls and jumping spiders have forward-facing eyes, providing accurate binocular vision over a relatively narrow field of view, whereas prey animals often have less acute all-round vision. Animals such as foxes can smell their prey even when it is concealed under 2 feet (60 cm) of snow or earth. Many predators have acute hearing, and some such as echolocating bats hunt exclusively by active or passive use of sound (Royal Saskatchewan Museum, 2012).
Predators including big cats, birds of prey, and ants share powerful jaws, sharp teeth, or claws which they use to seize and kill their prey. Some predators such as snakes and fish-eating birds like herons and cormorants swallow their prey whole; some snakes can unhinge their jaws to allow them to swallow large prey, while fish-eating birds have long spear-like beaks that they use to stab and grip fast-moving and slippery prey (Royal Saskatchewan Museum, 2012). Fish and other predators have developed the ability to crush or open the armored shells of molluscs (Vermeij, 1993).
Many predators are powerfully built and can catch and kill animals larger than themselves; this applies as much to small predators such as ants and shrews as to big and visibly muscular carnivores like the cougar and lion (Lafferty & Kuris, 2002; Getz, 2011; Royal Saskatchewan Museum, 2012).

Figure \(\PageIndex{10}\): a) Skull of brown bear has large pointed canines for killing prey, and self-sharpening carnassial teeth at rear for cutting flesh with a scissor-like action, b) Large compound eyes, sensitive antennae, and powerful jaws (mandibles) of jack jumper ant, c) Crab spider, an ambush predator with forward-facing eyes, catching another predator, a field digger wasp, d) Red-tailed hawk uses sharp hooked claws and beak to kill and tear up its prey, e) Specialist: a great blue heron with a speared fish, f) Indian python unhinges its jaw to swallow large prey like this chital.
Camouflage and mimicry


Figure \(\PageIndex{11}\): A camouflaged snow leopard in Ladakh (top photo). Striated frogfish uses camouflage and aggressive mimicry in the form of a fishing rod-like lure on its head to attract prey (bottom photo).
Camouflage is the use of any combination of materials, coloration, or illumination for concealment, either by making animals or objects hard to see, or by disguising them as something else. Predators may use camouflage to more effectively capture prey, while prey may use camouflage to more effectively evade predation. Members of the cat family such as the snow leopard (treeless highlands), tiger (grassy plains, reed swamps), ocelot(forest), fishing cat (waterside thickets), and lion (open plains) are camouflaged with coloration and disruptive patterns suiting their habitats (Cott, 1940).
In aggressive mimicry, certain predators, including insects and fishes, make use of coloration and behavior to attract prey. Female Photuris fireflies, for example, copy the light signals of other species, thereby attracting male fireflies, which they capture and eat (Lloyd, 1965). Flower mantises are ambush predators; camouflaged as flowers, such as orchids, they attract prey and seize it when it is close enough (Forbes, 2009). Frogfishes are extremely well camouflaged, and actively lure their prey to approach using an esca, a bait on the end of a rod-like appendage on the head, which they wave gently to mimic a small animal, gulping the prey in an extremely rapid movement when it is within range (Bester, 2017).
Other Adaptations
-
Venom - Many smaller predators such as the box jellyfish use venom to subdue their prey (Ruppert et al., 2005), and venom can also aid in digestion (as is the case for rattlesnakes and some spiders) (Cetaruk, 2005; Barceloux, 2008). The marbled sea snake that has adapted to egg predation has atrophied venom glands, and the gene for its three finger toxin contains a mutation (the deletion of two nucleotides) that inactives it. These changes are explained by the fact that its prey does not need to be subdued (Li et al., 2005).
-
Electric fields - Several groups of predatory fish have the ability to detect, track, and sometimes, as in the electric ray, to incapacitate their prey by generating electric fields using electric organs (Castello et al., 2009; Feulner et al., 2009; Catania, 2015). The electric organ is derived from modified nerve or muscle tissue (Kramer, 1996).
-
Physiology - Physiological adaptations to predation include the ability of predatory bacteria to digest the complex peptidoglycan polymer from the cell walls of the bacteria that they prey upon (Jurkevitch & Davidov, 2006). Carnivorous vertebrates of all five major classes (fishes, amphibians, reptiles, birds, and mammals) have lower relative rates of sugar to amino acid transport than either herbivores or omnivores, presumably because they acquire plenty of amino acids from the animal proteins in their diet (Karasov et al., 1988).
Predator/Prey Coevolution

Figure \(\PageIndex{12}\): Bats use echolocation to hunt moths at night.
To counter predation, prey have evolved defenses for use at each stage of an attack (Ruxton, 2004). They can try to avoid detection (Ruxton, 2004), such as by using camouflage and mimicry (Merilaita et al., 2017). They can detect predators (Caro, 2005) and warn others of their presence (Bergstrom & Lachmann, 2001; Getty, 2002). If detected, they can try to avoid being the target of an attack, for example, by signaling that they are toxic or unpalatable (Cott, 1940; Bowers et al., 1985), by forming groups (Beauchamp, 2012; Krause et al., 2002), or that a chase would be unprofitable (Ruxton et al., 2004; Caro, 2005). If they become a target, they can try to fend off the attack with defenses such as armor, quills, unpalatability, or mobbing (Dominey, 1983; Ruxton et al., 2004; Brodie, 2009) and they can escape an attack in progress by startling the predator (Cott, 1940; Merilaita et al., 2011; Edmunds, 2012), playing dead, shedding body parts such as tails, or simply fleeing (Caro, 2005).


Figure \(\PageIndex{13}\): Dead leaf mantis's camouflage makes it less visible to both predators and prey (top photo). Syrphid hoverfly misdirects predators by mimicking a wasp, but has no sting (bottom photo).
Predators and prey are natural enemies, and many of their adaptations seem designed to counter each other. For example, bats have sophisticated echolocation systems to detect insects and other prey, and insects have developed a variety of defenses including the ability to hear the echolocation calls (Jacobs & Bastian, 2017; Barbosa & Castellanos, 2005). Many pursuit predators that run on land, such as wolves, have evolved long limbs in response to the increased speed of their prey (Janis & Wilhelm, 1993). Their adaptations have been characterized as an evolutionary arms race, an example of the coevolution of two species (Dawkins & Krebs, 1979). In a gene centered view of evolution, the genes of predator and prey can be thought of as competing for the prey's body (Dawkins & Krebs, 1979). Though criticized, the "life-dinner" principle of Dawkins and Krebs predicts that this arms race is asymmetric: if a predator fails to catch its prey, it loses its dinner, while if it succeeds, the prey loses its life (Dawkins & Krebs, 1979).

Figure \(\PageIndex{14}\): Eastern coral snake, itself a predator, is venomous enough to kill predators that attack it, so when they avoid it, this behavior must be inherited, not learnt.
The metaphor of an arms race implies ever-escalating advances in attack and defense. However, these adaptations come with a cost; for instance, longer legs have an increased risk of breaking (Abrams, 1986), while the specialized tongue of the chameleon, with its ability to act like a projectile, is useless for lapping water, so the chameleon must drink dew off vegetation (Brodie, 1999).
16.1.3 Consequences of Predation
Predators affect their ecosystems not only directly by eating their own prey, but by indirect means such as reducing predation by other species, or altering the foraging behavior of an herbivore, as with the biodiversity effect of wolves on riverside vegetation or sea otters on kelp forests. This may explain population dynamics effects such as the cycles observed in lynx and snowshoe hares (Preisser et al., 2005; Peckarsky et al., 2008; Sheriff et al., 2020).
Biodiversity maintained by apex predation
Predators may increase the biodiversity of communities by preventing a single species from becoming dominant. Such predators are known as keystone species and may have a profound influence on the balance of organisms in a particular ecosystem (Bond, 2012). Introduction or removal of this predator, or changes in its population density, can have drastic cascading effects on the equilibrium of many other populations in the ecosystem. For example, grazers of a grassland may prevent a single dominant species from taking over (Bond, 2012).

Figure \(\PageIndex{15}\): Riparian willow recovery at Blacktail Creek, Yellowstone National Park, after reintroduction of wolves, the local keystone species and apex predator (Ripple & Beschta, 2004). Left, in 2002; right, in 2015
The elimination of wolves from Yellowstone National Park had profound impacts on the entire ecosystem’s structure and function. In Yellowstone, wolves are both keystone species and apex predators. Without predation, herbivores began to over-graze many woody browse species, affecting the area's plant populations. In addition, wolves often kept animals from grazing near streams, protecting the beavers' food sources. The removal of wolves had a direct effect on the beaver population, as their habitat became territory for grazing. Increased browsing on willows and conifers along Blacktail Creek due to a lack of predation caused channel incision because the reduced beaver population was no longer able to slow the water down and keep the soil in place. The predators were thus demonstrated to be of vital importance in the ecosystem (Ripple & Beschta, 2004).
Population dynamics
In the absence of predators, the population of a species can grow exponentially until it approaches the carrying capacity of the environment (Neal, 2004). Predators limit the growth of prey both by consuming them and by changing their behavior (Nelson et al., 2004). Increases or decreases in the prey population can also lead to increases or decreases in the number of predators, for example, through an increase in the number of young they bear.
Cyclical fluctuations have been seen in populations of predator and prey, often with offsets between the predator and prey cycles. A well-known example is that of the snowshoe hare and lynx. Over a broad span of boreal forests in Alaska and Canada, the hare populations fluctuate in near synchrony with a 10-year period, and the lynx populations fluctuate in response. This was first seen in historical records of animals caught by fur hunters for the Hudson Bay Company over more than a century (Krebs et al., 2001; Peckarsky et al., 2008; Krebs, 2014; BBC, 2015).
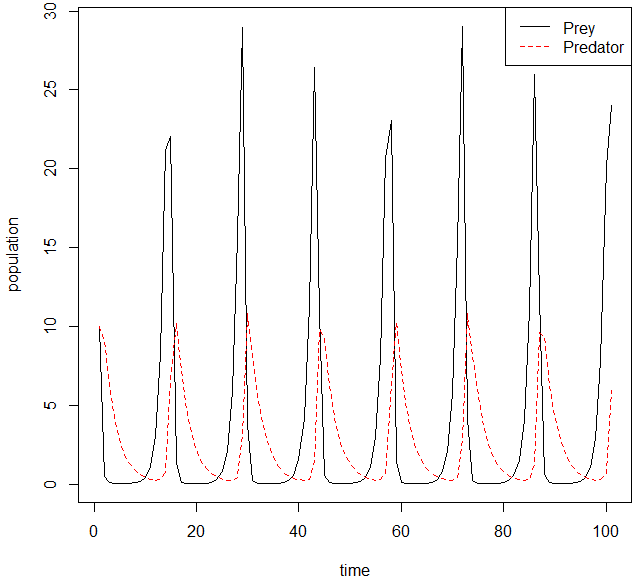
Figure \(\PageIndex{16}\): Predator-prey population cycles in a Lotka-Volterra model.
A simple model of a system with one species each of predator and prey, the Lotka–Volterra equations, predicts population cycles (Goel et al., 1971). However, attempts to reproduce the predictions of this model in the laboratory have often failed; for example, when the protozoan Didinium nasutum is added to a culture containing its prey, Paramecium caudatum, the latter is often driven to extinction (Levin et al., 2009).
The Lotka-Volterra equations rely on several simplifying assumptions, and they are structurally unstable, meaning that any change in the equations can stabilize or destabilize the dynamics (Murdoch et al., 2009; Nowak & May, 2000). For example, one assumption is that predators have a linear functional response to prey: the rate of kills increases in proportion to the rate of encounters. If this rate is limited by time spent handling each catch, then prey populations can reach densities above which predators cannot control them (Levin et al., 2009). Another assumption is that all prey individuals are identical. In reality, predators tend to select young, weak, and ill individuals, leaving prey populations able to regrow (Genovart et al., 2010).
Many factors can stabilize predator and prey populations (Rockwood, 2009). One example is the presence of multiple predators, particularly generalists that are attracted to a given prey species if it is abundant and look elsewhere if it is not (Rockwood, 2009). As a result, population cycles tend to be found in northern temperate and subarctic ecosystems because the food webs are simpler (Rockwood, 2009). The snowshoe hare-lynx system is subarctic, but even this involves other predators, including coyotes, goshawks and great horned owls, and the cycle is reinforced by variations in the food available to the hares (Rockwood, 2009).
A range of mathematical models have been developed by relaxing the assumptions made in the Lotka-Volterra model; these variously allow animals to have geographic distributions, or to migrate; to have differences between individuals, such as sexes and an age structure, so that only some individuals reproduce; to live in a varying environment, such as with changing seasons (Cushing, 2005; Thieme, 2003); and analyzing the interactions of more than just two species at once. Such models predict widely differing and often chaotic predator-prey population dynamics (Cushing, 2005; Kozlov & Vakulenko, 2013). The presence of refuge areas, where prey are safe from predators, may enable prey to maintain larger populations but may also destabilize the dynamics (Sih, 1987; McNair, 1986; Berryman et al., 2006; Ross & Garay, 2009).
Case study based on the CC-BY Paper By:
Glaspie, C. N., Seitz, R. D., & Lipcius, R. N. (2020). Are predator-prey model predictions supported by empirical data? Evidence for a storm-driven shift to an alternative stable state in a crab-clam system. Marine Ecology Progress Series, 645, 83-90.
Predators play a key role in ecosystem stability and function by consuming dominant competitors (Lubchenco & Gaines, 1981; Boudreau & Worm. 2012). Predators can also destabilize ecosystems or collapse food webs if they become too abundant (Estes et al., 2009), or if their prey do not have natural defenses against predation (Johnston et al., 2015). One of the ways the balance between predator and prey adaptations manifests itself in nature is through density-dependent predation. Predators can exhibit a numerical response to prey densities by increasing reproduction rates due to an overabundance of prey (demographic response) or by gathering in areas with relatively high densities of prey (aggregative response) (Holling, 1959). An individual predator may also adjust its predation rate to prey density through a ‘functional response’ (changes in a predator’s consumption rate in response to prey density).
Many mathematical models can be used to predict predator−prey dynamics (Briggs & Hoopes, 2004). These models contain nonlinear functions describing the density-dependent interactions between predator and prey. Due to nonlinearities, model behavior often includes shifts to alternative stable states (Drake & Griffen, 2010). The states may include extinction of one or both species, or coexistence steady states where both predator and prey are able to coexist at densities predicted by the model. The theory of alternative stable states predicts that ecosystems can exist under multiple "states" (sets of unique biotic and abiotic conditions). These alternative states are non-transitory and therefore considered stable over ecologically-relevant timescales. Ecosystems may transition from one stable state to another, in what is known as a state shift or regime shift, when perturbed. Due to ecological feedbacks, ecosystems display resistance to state shifts and therefore tend to remain in one state unless perturbations are large enough.
In their study, Glaspie et al. (2020) examined evidence for a storm-driven shift to a low-density state for the soft-shell clam Mya arenaria, which was once a biomass-dominant species in Chesapeake Bay, USA, in the face of predation by the blue crab Callinectes sapidus. Tropical Storm Agnes, which reached and remained in the Chesapeake Bay watershed from 21 to 23 June 1972, has long been suspected of causing long-term changes to the Bay (Orth & Moore, 1983). Tropical Storm Agnes was a ‘100-year storm’ that caused sustained, extremely low salinities and increased sedimentation throughout Chesapeake Bay (Schubel, 1976, Schubel et al., 1976). This storm has been blamed for accelerating the loss of seagrass Zostera marina (Orth & Moore, 1983) and oysters Crassostrea virginica (Haven et al. 1976) in Chesapeake Bay, although these losses were already in motion before the storm occurred. Unlike seagrass and oysters, declines in abundance of the soft-shell clam M. arenaria were uniquely related to the storm. M. arenaria was abundant enough to support a major commercial fishery throughout Chesapeake Bay prior to 1972 (Haven, 1970). Widespread mass mortality of M. arenaria occurred after the storm (Cory & Redding, 1976), and the fishery never recovered in lower Chesapeake Bay (Virginia) (Glaspie et al., 2018).
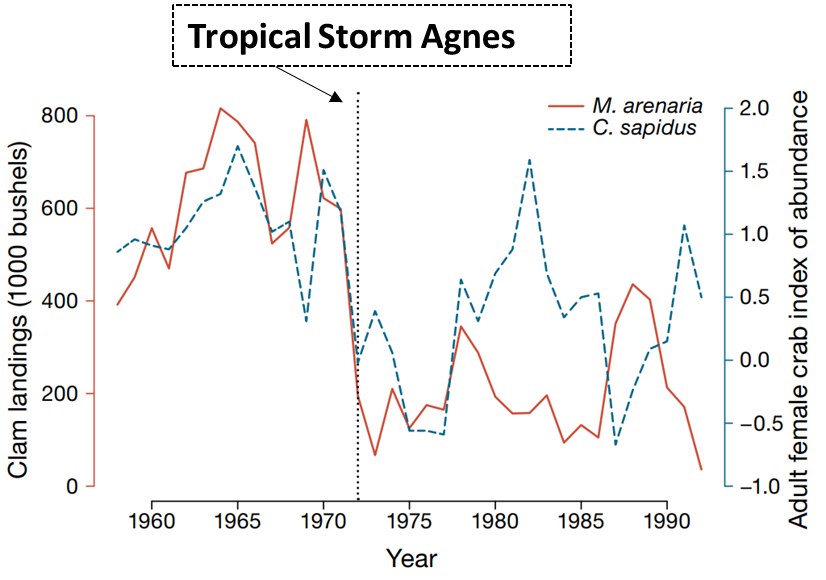
Figure \(\PageIndex{17}\): Though the boom-and-bust cycle of the species continued after the Tropical Storm Agnes, the storm clearly impacted the abundance of the populations and disrupted the pattern of their population cycles.
Using a 30-year dataset on soft-shell clams and blue crabs, Glaspie et al. (2020) found that Mya arenaria was subjected to a storm-driven shift to a low-density alternative stable state, which has been maintained by blue crab predation in Chesapeake Bay. Before the storm, clams were likely prey for juvenile crabs that entered the fishery at 1 yr of age, resulting in a positive correlation between crab and clam abundance with a lag of 1 yr (Figure \(\PageIndex{2a}\)). After the storm, crabs were likely consuming juvenile clams that would have been large enough to enter the fishery a year later, resulting in a negative correlation between clams and crab abundance with a lag of 1 yr (Figure \(\PageIndex{2b}\)). Both M. arenaria and Callinectes sapidus enter their respective fisheries after 1−1.5 yr (Newell & Hidu, 1986; Lipcius & van Engel, 1990), providing an explanation for the 1 yr lag. This is consistent with a shift from a system controlled from the bottom up (i.e., by the abundance of prey resources), to a system controlled from the top down by predation pressure on bivalves.

Figure \(\PageIndex{18}\): The change in trendlines from a positive correlation before the storm to a negative correlation after the storm again shows the impact natural disasters can have on the patterns of population cycles.
Sources
Abrams, P.A. (1986). Adaptive responses of predators to prey and prey to predators: The failure of the arms-race analogy. Evolution, 40(6), pp. 1229–1247. doi:10.1111/j.1558-5646.1986.tb05747.x. PMID 28563514. S2CID 27317468.
Barbosa, P., & Castellanos, I. (2005). Ecology of predator-prey interactions. Oxford University Press, p. 78. ISBN 9780199874545.
Barceloux, D.G. (2008). Medical toxicology of natural substances: Foods, fungi, medicinal herbs, plants, and venomous animals. Wiley, p. 1028. ISBN 978-0-470-33557-4.
Bar-Yam. Predator-prey relationships. New England Complex Systems Institute. Retrieved 7 September 2018.
Bengtson, S. (2002). Origins and early evolution of predation. In Kowalewski, M., & Kelley, P.H. (Eds.), The fossil record of predation. The Paleontological Society, pp. 289–317.
Bergstrom, C.T., & Lachmann, M. (2001). Alarm calls as costly signals of antipredator vigilance: The watchful babbler game. Animal Behaviour, 61(3), 535–543. CiteSeerX 10.1.1.28.773. doi:10.1006/anbe.2000.1636. S2CID 2295026.
Berryman, A.A., & Hawkins, Bradford A. (2006). The refuge as an integrating concept in ecology and evolution. Oikos, 115(1), 192–196. doi:10.1111/j.0030-1299.2006.15188.x.
Bester, C. (2017). Antennarius striatus. University of Florida. Retrieved 31 January 2018.
Bond, W.J. (2012). Keystone species. In Schulze, E.D., & Mooney, H.A. (Eds.), Biodiversity and Ecosystem Function. Springer. p. 237. ISBN 978-3642580017.
Boudreau, S.A., & Worm, B. (2012). Ecological role of large benthic decapods in marine ecosystems: A review. Mar. Ecol. Prog. Ser., 469, pp. 195−213
Bowers, M.D., Brown, I.L., & Wheye, D. (1985). Bird predation as a selective agent in a butterfly population. Evolution, 39(1), pp. 93–103. doi:10.1111/j.1558-5646.1985.tb04082.x. PMID 28563638. S2CID 12031679.
Bray, D. Eastern frogfish, Batrachomoeus dubius. Fishes of Australia. Archived from the original on 14 September 2014. Retrieved 14 September 2014.
Briggs, C.J., & Hoopes, M.F. (2004). Stabilizing effects in spatial parasitoid−host and predator−prey models: A review. Theor. Popul. Biol., 65, pp. 299−315
Brodie, E.D. (2009). Toxins and venoms. Current Biology, 19(20), R931–R935. doi:10.1016/j.cub.2009.08.011. PMID 19889364. S2CID 9744565.
Brodie, E.D. (1999). Predator-prey arms races. BioScience, 49(7), pp. 557–568. doi:10.2307/1313476. JSTOR 1313476.
Castello, M.E., Rodriguez-Cattaneo, A., Aguilera, P.A., Iribarne, L., Pereira, A.C., & Caputi, A.A. (2009). Waveform generation in the weakly electric fish Gymnotus coropinae (Hoedeman): The electric organ and the electric organ discharge. Journal of Experimental Biology, 212(9), pp. 1351–1364. doi:10.1242/jeb.022566. PMID 19376956.
Catania, K.C. (2015). Electric eels use high-voltage to track fast-moving prey. Nature Communications, 6(1), pp. 8638. Bibcode:2015NatCo...6.8638C. doi:10.1038/ncomms9638. ISSN 2041-1723. PMC 4667699. PMID 26485580.
Cetaruk, E.W. (2005). Rattlesnakes and other Crotalids. In Brent, J. (Eds.), Critical care toxicology: Diagnosis and management of the critically poisoned patient. Elsevier Health Sciences. p. 1075. ISBN 978-0-8151-4387-1.
Cory, R.L., & Redding, M.J. (1976). Mortalities caused by Tropical Storm Agnes to clams and oysters in the Rhode River area of Chesapeake Bay. In Anderson, A.M. (Eds.), The effects of Tropical Storm Agnes on the Chesapeake Bay estuarine system. The Johns Hopkins University Press, Baltimore, MD, p 478-487.
Cott. (1940). pp. 368–389
Cougar. Hinterland Who's Who. Canadian Wildlife Service and Canadian Wildlife Federation. Archived from the original on 18 May 2007. Retrieved 22 May 2007.
Cushing, J.M. (2005). Book reviews | Mathematics in population biology, by Horst R. Thiene. Bulletin of the American Mathematical Society, 42(4), pp. 501–505. doi:10.1090/S0273-0979-05-01055-4.
Dawkins, R., & Krebs, J.R. (1979). Arms races between and within species. Proceedings of the Royal Society B: Biological Sciences, 205(1161), pp. 489–511. Bibcode:1979RSPSB.205..489D. doi:10.1098/rspb.1979.0081. PMID 42057. S2CID 9695900.
deVries, M.S., Murphy, E.A.K., & Patek S.N. (2012). Strike mechanics of an ambush predator: The spearing mantis shrimp. Journal of Experimental Biology, 215(Pt 24), pp. 4374–4384. doi:10.1242/jeb.075317. PMID 23175528.
Dominey, W.J. (1983). Mobbing in colonially nesting fishes, especially the bluegill, Lepomis macrochirus. Copeia, (4), pp. 1086–1088. doi:10.2307/1445113. JSTOR 1445113.
Drake, J.M., & Griffen, B.D. (2010). Early warning signals of extinction in deteriorating environments. Nature, 467, pp. 456−459
Eastman, L.B., & Thiel, M. (2015). Foraging behavior of crustacean predators and scavengers. In Thiel, M., & Watling, L. (Eds.), Lifestyles and feeding biology. Oxford University Press. pp. 535–556. ISBN 9780199797066.
Edmunds, M. (2012). Deimatic Behavior. Springer. Retrieved 31 December 2012.
Estes, J.A., Doak, D.F., Springer, A.M., & Williams, T.M. (2009). Causes and consequences of marine mammal population declines in southwest Alaska: A food-web perspective. Philos. Trans. R. Soc. B., 364, pp. 1647−1658
Feulner, P. G., Plath, M., Engelmann, J., Kirschbaum, F., & Tiedemann, R. (2009). Electrifying love: Electric fish use species-specific discharge for mate recognition. Biology Letters, 5(2), pp. 225–228. doi:10.1098/rsbl.2008.0566. PMC 2665802. PMID 19033131.
Forbes, L.S. (1989). Prey defenses and predator handling behaviour: The dangerous prey hypothesis. Oikos, 55(2), pp. 155–158. doi:10.2307/3565418. JSTOR 3565418.
Forbes, P. (2009). Dazzled and deceived: Mimicry and camouflage. Yale University Press, p. 134. ISBN 978-0-300-17896-8.
Gazda, S.K., Connor, R.C., Edgar, R.K., & Cox, F. (2005). A division of labour with role specialization in group-hunting bottlenose dolphins (Tursiops truncatus) off Cedar Key, Florida. Proceedings of the Royal Society, 272(1559), pp. 135–140. doi:10.1098/rspb.2004.2937. PMC 1634948. PMID 15695203.
Genovart, M., Negre, N., Tavecchia, G., Bistuer, A., Parpal, L., & Oro, D. (2010). The young, the weak and the sick: Evidence of natural selection by predation. PLOS ONE, 5(3): e9774. Bibcode:2010PLoSO...5.9774G. doi:10.1371/journal.pone.0009774. PMC 2841644. PMID 20333305.
Getty, T. (2002). The discriminating babbler meets the optimal diet hawk. Anim. Behav., 63(2), pp. 397–402. doi:10.1006/anbe.2001.1890. S2CID 53164940.
Glaspie, C.N., Seitz, R.D., Ogburn, M.B., Dungan, C.F., & Hines, A.H. (2018). Impacts of habitat, predators, recruitment, and disease on soft-shell clams Mya arenaria and stout razor clams Tagelus plebeius in Chesapeake Bay. Mar. Ecol. Prog. Ser., 603, pp. 117−133
Goel, N.S., Maitra, S.C., & Montroll, E.W. (1971). On the Volterra and other non-linear models of interacting populations. Academic Press. ISBN 978-0122874505.
Goldbogen, J.A., Calambokidis, J., Shadwick, R.E., Oleson, E.M., McDonald, M.A., & Hildebrand, J.A. (2006). Kinematics of foraging dives and lunge-feeding in fin whales. Journal of Experimental Biology, 209(7), pp. 1231–1244. doi:10.1242/jeb.02135. PMID 16547295. S2CID 17923052. Archived from the original (PDF) on 13 August 2011. Retrieved 4 September 2018.
Gremillet, D., Wilson, R.P., Wanless, S., & Chater, T. (2000). Black-browed albatrosses, international fisheries and the Patagonian Shelf. Marine Ecology Progress Series, 195, pp. 69–280. Bibcode:2000MEPS..195..269G. doi:10.3354/meps195269.
Griffiths, D. (1980). Foraging costs and relative prey size. The American Naturalist, 116(5), pp. 743–752. doi:10.1086/283666. JSTOR 2460632. S2CID 85094710.
Gurr, G.M., Wratten, S.D., & Snyder, W.E. (2012). Biodiversity and insect pests: Key issues for sustainable management. John Wiley & Sons, pp. 105. ISBN 978-1-118-23185-2.
Haven, D.S. (1970). A study of the hard and soft clam resources of Virginia: Annual contract report for the period 1 July 1969 through 30 June 1970. Virginia Institute of Marine Science, Gloucester Point, VA.
Haven, D.S., Hargis, W.J., Loesch, J.G., & Whitcomb, J.P. (1976). The effect of Tropical Storm Agnes on oysters, hard clams, soft clams, and oyster drills in Virginia. In Anderson, A.M. (Eds.), The effects of Tropical Storm Agnes on the Chesapeake Bay estuarine system. The Johns Hopkins University Press, Baltimore, MD, pp. 488−508.
Holling, C. (1959). The components of predation as revealed by a study of small mammal predation of the European pine sawfly. Can. Entomol., 91, pp. 293−320
Hubel, T.Y., Myatt, J.P., Jordan, N.R., Dewhirst, O.P., McNutt, J.W., & Wilson, A.M. (2016). Energy cost and return for hunting in African wild dogs. Nature Communications, 7: 11034. doi:10.1038/ncomms11034. PMC 4820543. PMID 27023457.
Janis, C.M., & Wilhelm, P.B. (1993). Were there mammalian pursuit predators in the Tertiary? Dances with wolf avatars. Journal of Mammalian Evolution, 1(2), pp. 103–125. doi:10.1007/bf01041590. S2CID 22739360.
Jurkevitch, E., & Davidov, Y. (2006). Phylogenetic diversity and evolution of predatory prokaryotes. Predatory Prokaryotes. Springer. pp. 11–56. doi:10.1007/7171_052. ISBN 978-3-540-38577-6.
Kane, A., Healy, K., Guillerme, T., Ruxton, G.D., & Jackson, A.L. (2017). A recipe for scavenging in vertebrates – the natural history of a behaviour. Ecography, 40(2), pp. 324–334. doi:10.1111/ecog.02817
Karasov, W.H., & Diamond, J.M. (1988). Interplay between physiology and ecology in digestion. BioScience, 38(9), pp. 602–611. doi:10.2307/1310825. JSTOR 1310825.
Kozlov, V., & Vakulenko, S. (2013). On chaos in Lotka–Volterra systems: An analytical approach. Nonlinearity, 26(8), pp. 2299–2314. Bibcode:2013Nonli..26.2299K. doi:10.1088/0951-7715/26/8/2299.
Kramer, D.L. (2001). Foraging behavior. In Fox, C.W., Roff, D.A., & Fairbairn, D.J. (Eds.), Evolutionary ecology: Concepts and case studies. Oxford University Press, pp. 232–238. ISBN 9780198030133. Archived from the original (PDF) on 12 July 2018. Retrieved 20 September 2018.
Krause, J., & Ruxton, G.D. (2002). Living in groups. Oxford University Press, pp. 13–15. ISBN 9780198508182.
Krebs, C.J., Boonstra, R., Boutin, S., & Sinclair, A.R.E. (2001). What drives the 10-year cycle of snowshoe hares?. BioScience, 51(1), pp. 25. doi:10.1641/0006-3568(2001)051[0025:WDTYCO]2.0.CO;2.
Krebs, C., & Myers, J. (2014). The snowshoe hare 10-year cycle – a cautionary tale. Ecological rants. University of British Columbia. Retrieved 2 October 2018.
Kruuk, H. (1972). The spotted hyena: A study of predation and social behaviour. University of California Press, pp. 107–108. ISBN 978-0226455082.
Lafferty, K.D., & Kuris, A.M. (2002). Trophic strategies, animal diversity and body size. Trends in Ecology and Evolution, 17(11), pp. 507–513. doi:10.1016/s0169-5347(02)02615-0. Archived from the original (PDF) on 3 October 2019.
Levin, S.A., Carpenter, S.R., Godfray, H.C.J., Kinzig, A.P., Loreau, M., Losos, J.B., Walker, B., & Wilcove, D.S. (2009). The Princeton guide to ecology. Princeton University Press, pp. 204–209. ISBN 9781400833023.
Li, M., Fry, B.G., & Kini, R.M. (2005). Eggs-only diet: Its implications for the toxin profile changes and ecology of the marbled sea snake (Aipysurus eydouxii). Journal of Molecular Evolution, 60(1), pp. 81–89. Bibcode:2005JMolE..60...81L. doi:10.1007/s00239-004-0138-0. PMID 15696370. S2CID 17572816.
Lipcius, R.N., & van Engel, W.A. (1990). Blue crab population dynamics in Chesapeake Bay: Variation in abundance (York River 1972−1988) and stock−recruit functions. Bull Mar. Sci., 46, pp. 180−194
Lloyd J.E. (1965). Aggressive mimicry in Photuris: Firefly femmes fatales. Science, 149(3684), pp. 653–654. Bibcode:1965Sci...149..653L. doi:10.1126/science.149.3684.653. PMID 17747574. S2CID 39386614.
Lubchenco, J., & Gaines, S.D. (1981). A unified approach to marine plant−herbivore interactions. I. Populations and communities. Annu. Rev. Ecol. Syst., 12, pp. 405−437
MacArthur, R.H. (1984). The economics of consumer choice. Geographical ecology: Patterns in the distribution of species. Princeton University Press, pp. 59–76. ISBN 9780691023823.
McNair, J.N. (1986). The effects of refuges on predator-prey interactions: A reconsideration. Theoretical Population Biology, 29,(1), pp. 38–63. doi:10.1016/0040-5809(86)90004-3. PMID 3961711.
Merilaita, S., Scott-Samuel, N.E., & Cuthill, I.C. (2017). How camouflage works. Philosophical Transactions of the Royal Society B: Biological Sciences, 372(1724): 20160341. doi:10.1098/rstb.2016.0341. PMC 5444062. PMID 28533458.
Merilaita, S., Vallin, A., Kodandaramaiah, U., Dimitrova, M., Ruuskanen, S., & Laaksonen, T. (2011). Number of eyespots and their intimidating effect on naïve predators in the peacock butterfly. Behavioral Ecology, 22(6), pp. 1326–1331. doi:10.1093/beheco/arr135.
Moore, T.Y., & Biewener, A.A. (2015). Outrun or outmaneuver: Predator–prey interactions as a model system for integrating biomechanical studies in a broader ecological and evolutionary context. Integrative and Comparative Biology, 55(6), pp. 1188–97. doi:10.1093/icb/icv074. PMID 26117833.
Neal, D. (2004). Introduction to population biology. Cambridge University Press, pp. 68–69. ISBN 9780521532235.
Newell, C.R., & Hidu, H. (1986). Species profiles: Life histories and environmental requirements of coastal fishes and invertebrates (North Atlantic)—softshell clam. US Fish Wildl. Serv. Biol. Rep., 82(11.53). US Army Corps of Engineers, TR EL-82-4. https: //apps.dtic.mil/dtic/tr/fulltext/ u2/ a174837. Pdf
Nowak, M., & May, R.M. (2000). Virus dynamics: Mathematical principles of immunology and virology. Oxford University Press, p. 8. ISBN 9780191588518.
Orth, R.J., & Moore, K.A. (1983). Chesapeake Bay: An unprecedented decline in submerged aquatic vegetation. Science, 222, pp. 51−53
Peckarsky, B.L., Abrams, P.A., Bolnick, D.I., Dill, L.M., Grabowski, J.H., Luttbeg, B., Orrock, J.L., Peacor, S.D., Preisser, E.L., Schmitz, O.J., & Trussell, G.C. (2008). Revisiting the classics: Considering nonconsumptive effects in textbook examples of predator–prey interactions. Ecology, 89(9), pp. 2416–2425. doi:10.1890/07-1131.1. PMID 18831163.
Perry, G. (1999). The evolution of search modes: Ecological versus phylogenetic perspectives. The American Naturalist, 153(1), pp. 98–109. doi:10.1086/303145. PMID 29578765. S2CID 4334462.
Pianka, E.R. (2011). Evolutionary ecology (7th (eBook) ed.). pp. 78–83.
Pikes (Esocidae). Indiana Division of Fish and Wildlife. Retrieved 3 September 2018.
Predator & prey: Adaptations. (2012). Royal Saskatchewan Museum. Archived from the original (PDF) on 3 April 2018. Retrieved 19 April 2018.
Predators and their prey. BBC. Retrieved 7 October 2015.
Poulin, R. (2011). The many roads to parasitism: A tale of convergence. In Rollinson, D., & Hay, S.I. (Eds.), Advances in parasitology, 74. Academic Press. pp. 27–28. doi:10.1016/B978-0-12-385897-9.00001-X. ISBN 978-0-12-385897-9. PMID 21295676.
Poulin, R., & Randhawa, H.S. (2015). Evolution of parasitism along convergent lines: From ecology to genomics. Parasitology, 142(Suppl 1): S6–S15. doi:10.1017/S0031182013001674. PMC 4413784. PMID 24229807.
Preisser, E.L., Bolnick, D.I., & Benard, M.F. (2005). Scared to death? The effects of intimidation and consumption in predator–prey interactions. Ecology, 86(2), pp. 501–509. doi:10.1890/04-0719. ISSN 0012-9658.
Ruppert, E.E., Fox, R.S., & Barnes, R.D. (2004). Invertebrate zoology, 7th edition. Cengage Learning. pp. 153–154. ISBN 978-81-315-0104-7.
Ruxton, Sherratt & Speed 2004, pp. vii–xii
Sanders, J.G., Beichman, A.C., Roman, J., Scott, J.J., Emerson, D., McCarthy, J.J., & Girguis, P.R. (2015). Baleen whales host a unique gut microbiome with similarities to both carnivores and herbivores. Nature Communications, 6, pp. 8285. Bibcode:2015NatCo...6.8285S. doi:10.1038/ncomms9285. PMC 4595633. PMID 26393325.
Schmidt, J.O. (2009). Wasps. ScienceDirect. Encyclopedia of Insects (Second ed.). pp. 1049–1052. doi:10.1016/B978-0-12-374144-8.00275-7
Schubel, J.R. (1976). Effects of Agnes on the suspended sediment on the Chesapeake Bay and contiguous shelf waters. In Schubel, J.R. (Eds.), The effects of Tropical Storm Agnes on the Chesapeake Bay estuarine system. The Johns Hopkins University Press, Baltimore, MD, p 179−200
Schubel, J.R., Carter, H.H., & Cronin, W.B. (1976). Effects of Agnes on the distribution of salinity along the main axis of the Bay and in contiguous shelf waters. In Ruzecki, E.P. (Eds.), The effects of Tropical Storm Agnes on the Chesapeake Bay estuarine system. The Johns Hopkins University Press, Baltimore, MD, p 33−65
Sheriff, M.J., Peacor, S.D., Hawlena, D., Thaker, M., & Gaillard, J.M. (2020). Non-consumptive predator effects on prey population size: A dearth of evidence. Journal of Animal Ecology, 89(6), pp. 1302–1316. doi:10.1111/1365-2656.13213. PMID 32215909.
Sih, A. (1987). Prey refuges and predator-prey stability. Theoretical Population Biology, 31, pp. 1–12. doi:10.1016/0040-5809(87)90019-0.
Stevens, A.N.P. (2010). Predation, herbivory, and parasitism. Nature Education Knowledge, 3(10), pp. 36. Retrieved 12 February 2018.
Trapdoor spider. (2014). Arizona-Sonora Desert Museum. Retrieved 12 December 2014.
Trapdoor spiders. BBC. Retrieved 12 December 2014.
Thieme, H.R. (2003). Mathematics in population biology. Princeton University Press. ISBN 978-0-691-09291-1.
Tyus, H.M. (2011). Ecology and conservation of fishes. CRC Press, p. 233. ISBN 978-1-4398-9759-1.
Vermeij, G.J. (1993). Evolution and escalation: An ecological history of life. Princeton University Press, pp. 11 and passim. ISBN 978-0-691-00080-0.
Wetzel, R.G., & Likens, G.E. (2000). Predator-prey interactions. Limnological Analyses. Springer. pp. 257–262. doi:10.1007/978-1-4757-3250-4_17. ISBN 978-1-4419-3186-3.
Williams, A.C., & Flaxman, S.M. (2012). Can predators assess the quality of their prey's resource? Animal Behaviour, 83(4), pp, 883–890. doi:10.1016/j.anbehav.2012.01.008. S2CID 53172079.