
19.1.13: Vertebrates
- Page ID
- 5965

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Chordata
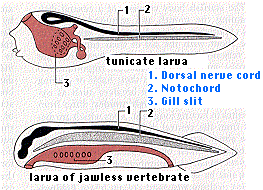
During their embryonic development, all chordates pass through a stage called the pharyngula with these features:
- The cephalochordates and tunicates never develop a vertebral column. They are thus "invertebrates" and are discussed with the other invertebrates.
- Craniata The vast majority of chordates have a skull enclosing their brain, eyes, inner ear, etc.). All but one group of these (the hagfishes) also convert their notochord into a vertebral column or backbone thus qualifying as vertebrates.
Vertebrata
Although hagfishes, never replace their notochord with a vertebral column, and thus might seem not to qualify as vertebrates, they share a number of other features with other vertebrates and certainly should be classified with them. Still uncertain is whether they represent the most primitive vertebrates or are simply degenerate vertebrates (probably the latter).
All the other members of the craniata convert their notochord into a vertebral column or "backbone" (even though in some it is made of cartilage not bone). They also differ from all other animals in having quadrupled their HOX gene cluster; that is, they have 4 different clusters of HOX genes (on 4 separate chromosomes). Perhaps this acquisition played a key role in the evolutionary diversity that so characterizes the vertebrates.
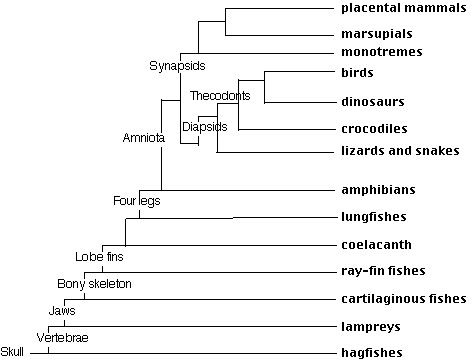
The vertebrates are subdivided into the
- jawless vertebrates (Agnatha)
- jawed vertebrates (Gnathostomata)
Agnatha
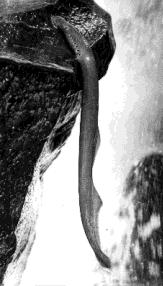
Lampreys and hagfishes are the only jawless vertebrates to survive today. They both have a round mouth and for this reason are often referred to as cyclostomes. They are the most primitive of the vertebrates. By "primitive", a biologist means that they are the least changed from the first vertebrates. Besides lacking jaws,
- They have no paired pectoral (shoulder) or pelvic (hip) fins.
- Their notochord persists for life, never being completely replaced by a backbone even in the lampreys.
- They have no scales.
- The axons of their neurons are unmyelinated (like those of all invertebrates).
- Lampreys have both an innate immune system and an adaptive immune system, but the latter is entirely different from that found in the jawed vertebrates.
The photo (courtesy of the Carolina Biological Supply Company) is of the West Coast lamprey. Note the gill slits and the absence of paired pectoral and pelvic fins.
Gnathostomata
As well as having jaws, all the members of this group have
- Myelin sheaths around the axons of their neurons. This permits much more rapid transmission of nerve impulses - a trait probably as important for active vertebrates as their jaws.
- An adaptive immune system backing up their innate immune system.
Cartilaginous Fishes (Chondrichthyes)
Fossils of cartilaginous fishes become abundant in deposits dating to the Devonian period. They were very much like the sharks of today. The group, which today is made up of some 1,188 species of sharks, skates and rays gets its name from the fact that their skeleton is made of cartilage, not bone.
With their gills exposed to sea water, all marine fishes are faced with the problem of conserving body water in a strongly hypertonic environment. Sea water is about 3.5% salt, over 3 times that of vertebrate blood. The cartilaginous fishes solve the problem by maintaining such a high concentration of urea in their blood (2.5% — far higher than the ~0.02% of other vertebrates) that it is in osmotic balance with - that is, is isotonic to - sea water.
This ability develops late in embryology, so the eggs of these species cannot simply be released in the sea. Two solutions are used:
- Enclose the egg in an impervious case filled with isotonic fluid before depositing it in the sea.
- Retain the eggs and embryos within the mother's body until they are capable of coping with the marine environment.
Both these solutions require internal fertilization, and the cartilaginous fishes were the first vertebrates to develop this. The pelvic fins of the male are modified for depositing sperm in the reproductive tract of the female.
Bony Fishes (Osteichthyes)
As their name indicates, the skeleton in this group is made of bone. The group is subdivided into the
- ray-finned fishes (Actinopterygii)
- lobe-finned fishes (Sarcopterygii)
Ray-finned fishes
- Their fins are thin and supported by spines.
- There are over 30,000 species (representing more than half of all living vertebrates).
- They are an important part of the human diet in many areas of the world and, in affluent nations, support a large sports fishing industry.
Although the earliest bony fishes may have appeared late in the Silurian period, their fossils become abundant in freshwater deposits of the Devonian period. In addition to gills, these fishes had a pair of pouched outgrowths from the pharynx which served as lungs. They were inflated with air taken in through the mouth and may have provided a backup gas exchange organ when the water became too warm and stagnant to carry enough dissolved oxygen. Their kidneys were adapted for the hypotonic environment in which they lived.
These animals diversified through the remainder of the Devonian period (which is often called the "Age of Fishes"). Some migrated to the oceans. In this more stable environment, their lungs became transformed into a swim bladder with which they could alter buoyancy. Their kidneys became transformed as well adapting them to their new - hypertonic - surroundings.
Lobe-finned fishes
The only ones to survive today are:
- two species of coelacanths. Coelacanths were long thought to have become extinct at the end of the Mesozoic era, some 70 million years ago. But in December 1938, a living coelacanth, Latimeria chalumnae, was pulled up from the depths of the ocean off the east coast of Africa. Since then, over 200 additional specimens have been caught.
- several species of lungfish found in Africa, South America, and Australia.
The nostrils of bony fishes open only to the outside and are used for smelling. Some of the lobe-finned fishes developed internal openings to their nostrils. This made it possible to breath air with the mouth closed as modern lungfishes do.
Judging from present-day lungfishes, two other significant adaptations evolved in this group:
- two atria and a partial septum in the ventricle of the heart (similar to the frog heart). This permitted a partial separation of oxygenated blood returning from the lung(s) and the deoxygenated blood returning from the rest of the body.
- an enzyme system to convert ammonia into the less toxic urea. This mechanism is highly-developed in the African and South American lungfishes. While in the water, these fishes excrete their waste nitrogen as ammonia, just as most ray-finned fishes do. In time of drought, these animals burrow in the mud and switch to urea production.
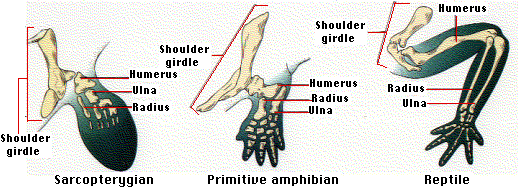
With their bony limbs and lungs inherited from their lobe-finned ancestors, amphibians were so successful during the Carboniferous (Mississippian and Pennsylvanian periods) that these periods are known as the Age of Amphibians.
The Carboniferous was followed by the Permian, when the earth became colder and dryer. The fortunes of the amphibians began to decline until only three groups - totaling about 6500 species - remain today:
- frogs and toads (Anura) (The one pictured is Rana pipiens, the leopard frog.)
- salamanders and newts (Urodela)
- caecilians (Apoda), which are rare, limbless, tropical animals.
As the name suggests, amphibians are only semiterrestrial:
- Their skin is soft and moist so they are at risk of desiccation in dry surroundings.
- Their eggs have no waterproof covering so
- they must be laid in water (which makes them useful animals for studying embryonic development) where they are fertilized or
- placed within the mother's body (some use a pouch in the skin, some use their mouth, some even use their stomach — which stops secreting acid and enzymes for the duration!) after external fertilization.
Amniotes (Amniota)
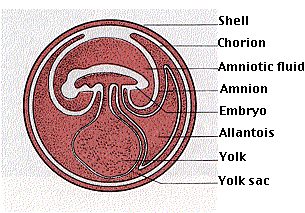
Some 310 million years ago (in the Pennsylvanian), some amphibians evolved the ability to lay shelled, yolk-filled eggs. The embryo developing within the egg produces 4 extraembryonic membranes:
- amnion, which surrounds the embryo with a fluid as watery as the pond water around a frog's egg (and accounts for the name amniota)
- chorion, which serves for gas exchange
- allantois, which serves both for gas exchange and to store metabolic wastes
- yolk sac, which supplies the embryo with food

With the arrival of the cold, dry Permian, reptiles were well-adapted to survive because of their development of a shelled, yolk-filled egg which could be deposited on land without danger of drying out. The photo (courtesy of the Carolina Biological Supply Company) shows an American chameleon emerging from its egg.
Other adaptations that enabled the reptiles to flourish for the next 220 million years were:
- a dry, water-impermeable skin
- lungs inflated by expansion of the rib cage
- a partial septum in the ventricle reducing the mixing of oxygenated and deoxygenated blood
Beginning late in the Paleozoic era and exploding in the Triassic period, the reptiles underwent a remarkable adaptive radiation producing the diapsids. This group developed the ability to convert their nitrogenous waste into uric acid. Uric acid is almost insoluble in water so its excretion involves little loss of water. (It is the whitish paste that pigeons leave on statues.) This modification largely freed the diapsids and their descendants from a dependence on drinking water; the water in their food is usually sufficient.
Diapsid evolution soon produced:
- lizards and snakes (Squamata - some 6,300 species survive today);
- turtles
- thecodonts.
Thecodonts were able to run fast by rising up on their hind legs, which became larger than their front legs, and using their long tail for balance. The group diversified into:
- crocodiles and alligators (Crocodilia — 22 species survive today)
- an extraordinary array of dinosaurs from some of which evolved today's birds.
Feathers are the feature that most clearly distinguishes the birds from their dinosaur ancestors. These scaly skin outgrowths provide a light, strong surface for the wings;
- heat insulation, making it possible to be small but still warm-blooded.
Other adaptations are:All of these adaptations help birds to fly (to escape predators and find suitable food and nesting sites). Almost 10,000 species are known today.