
16.3D: Angiosperm Life Cycle
- Page ID
- 5791

Angiosperms are the flowering plants (today the most abundant and diverse plants on earth). Most are terrestrial and all lack locomotion. This poses several problems. Gametes are delicate single cells. For two plants to cross fertilize, there must be a mechanism for the two gametes to reach each other safely. Moreover, there must also be a mechanism to disperse their offspring far enough away from the parent so that they do not have to compete with the parent for light, water, and soil minerals. The functions of the flower solve both of these problems.
The Flower and Its Pollination
In angiosperms, meiosis in the sporophyte generation produces two kinds of spores: (1) microspores which develop in the microsporangium and will germinate and develop into the male gametophyte generation and (2) megaspores that develop in the megasporangium will develop into the female gametophyte generation. Both types of sporangia are formed in flowers. In most angiosperms, the flowers are perfect: each has both microsporangia and megasporangia, although some angiosperms are imperfect, having either microsporangia or megasporangia but not both.
- Monoecious plants have both types of imperfect flower on the same plant.
- Dioecious plants have imperfect flowers on separate plants; that is, some plants are male, some female. Examples include willows, poplars, and the date palm. Most dioecious plants use an X-Y system of of sex determination like that in mammals. However, a few species use an X-to-autosome ratio system like that of Drosophila, and a very few use a ZW system like that of birds and lepidoptera.
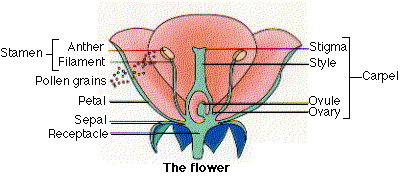
Flowers develop from flower buds. Each bud contains 4 concentric whorls of tissue. From the outer to the inner, these develop into
- a whorl of sepals (collectively called the calyx)
- a whorl of petals (collectively called the corolla)
- stamens in which the microsporangia form
- carpels in which the megasporangia form.
Stamens
Each stamen consists of a lobed anther, containing the microsporangia and supported by a thin filament. Meiosis of the diploid microspore mother cells in the anther produces four haploid microspores. Each of these develops into a pollen grain consisting of a larger vegetative cell (also called the tube cell) inside of which is a a smaller germ cell (also called the generative cell). At some point, depending on the species, the germ cell divides by mitosis to produce 2 sperm cells.
Carpels
Carpels consist of a stigma, usually mounted at the tip of a style with an ovary at the base. Often the entire whorl of carpels is fused into a single pistil. The megasporangia, called ovules, develop within the ovary. Meiosis of the megaspore mother cell in each ovule produces 4 haploid cells, a large megaspore and 3 small cells that disintegrate.
Development of the megaspore
The nucleus of the megaspore undergoes three successive mitotic divisions. The 8 nuclei that result are distributed and partitioned off by cell walls to form the embryo sac. This is the mature female gametophyte generation. The egg cell will start the new sporophyte generation if it is fertilized. It is flanked by 2 synergid cells. In several (perhaps all) angiosperms, they secrete an attractant that guides the pollen tube through the micropyle into the embryo sac. The large central cell, which in most angiosperms contains 2 polar nuclei, will after its fertilization develop into the endosperm of the seed. It also contains 3 antipodal cells.
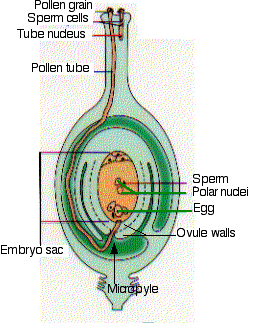
Pollination
When a pollen grain reaches the stigma, it germinates into a pollen tube. If it hasn't done so already, the germ cell divides by mitosis forming 2 sperm cells. These, along with the tube nucleus (also known as the vegetative nucleus), migrate down the pollen tube as it grows through the style, the micropyle, and into the ovule chamber.
In Arabidopsis the pollen tube follows a gradient of increasing concentration of a small defensin-like protein secreted by the synergids. The pollen tube with its contents makes up the mature male gametophyte generation.
Double fertilization
The pollen tube enters the ovule through the micropyle and ruptures. One sperm cell fuses with the egg forming the diploid zygote. The other sperm cell fuses with the polar nuclei forming the endosperm nucleus. Most angiosperms have two polar nuclei so the endosperm is triploid (3n). The tube nucleus disintegrates.
Most angiosperms have mechanisms by which they avoid self-fertilization.
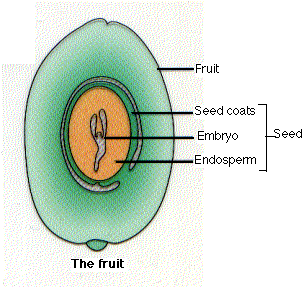
Seeds
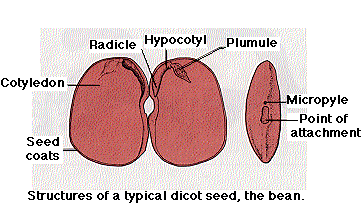
After double fertilization, each ovule develops into a seed, which consists of
- a plumule, made up of two embryonic leaves, which will become the first true leaves of the seedling, and a terminal (apical) bud. The terminal bud contains the meristem at which later growth of the stem takes place.
- One or two cotyledons which store food that will be used by the germinating seedling. Angiosperms that produce seeds with two cotyledons are called dicots. Examples include beans, squashes, Arabidopsis. Angiosperms whose seeds contain only a single cotyledon are monocots. Examples include corn and other grasses.
- The hypocotyl and radicle, which will grow into the part of the stem below the first node ("hypocotyl" = below the cotyledons) and primary root respectively. The development of each of the parts of the plant embryo depends on gradients of the plant hormone, auxin.
- In addition to the embryo plant (derived from the zygote), each seed is covered with protective seed coats derived from the walls of the ovule.
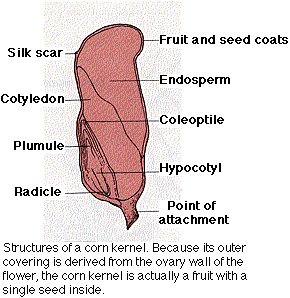
The food in the cotyledons is derived from the endosperm which, in turn, received it from the parent sporophyte. In many angiosperms (e.g., beans), when the seeds are mature, the endosperm has been totally consumed and its food transferred to the cotyledons. In others (some dicots and all monocots), the endosperm persists in the mature seed.
The seed is thus a dormant embryo sporophyte with stored food and protective coats. Its two functions are
- dispersal of the species to new locations (aided in angiosperms by the fruit)
- survival of the species during unfavorable climatic periods (e.g., winter). "Annual" plants (e.g., beans, cereal grains, many weeds) can survive freezing only as seeds. When the parents die in the fall, the seeds remain alive — though dormant— over the winter. When conditions are once more favorable, germination occurs and a new generation of plants develops.
Fruits
Fruits are a development of the ovary wall and sometimes other flower parts as well. As seeds mature, they release the hormone auxin, which stimulates the wall of the ovary to develop into the fruit. In fact, commercial fruit growers may stimulate fruit development in unpollinated flowers by applying synthetic auxin to the flower.
Fruits promote the dispersal of their content of seeds in a variety of ways.
- Wind. The maple "key" and dandelion parachute are examples.
- Water. Many aquatic angiosperms and shore dwellers (e.g., the coconut palm) have floating fruits that are carried by water currents to new locations.
- Hitchhikers. The cocklebur and sticktights achieve dispersal of their seeds by sticking to the coat (or clothing) of a passing animal.
- Edible fruits. Nuts and berries entice animals to eat them. Buried and forgotten (nuts) or passing through their g.i. tract unharmed (berries), the seeds may end up some distance away from the parent plant.
- Mechanical. Some fruits, as they dry, open explosively expelling their seeds. The pods of many legumes (e.g., wisteria) do this.