
15.5B: Vertebrate Kidneys
- Page ID
- 5461

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)All vertebrates have kidneys. Like the human kidney, they are made up of many nephrons. However, there are differences in the structure and functioning of various vertebrate kidneys that adapt them to the environment in which the animals live.
Freshwater Vertebrates
All animals that live in fresh water must cope with a continual inflow of water from their hypotonic environment. In order to maintain homeostasis of its extracellular fluid (ECF), the freshwater fish must excrete this excess water. Contraction of its heart (powered by ATP) provides the pressure to force the water, small molecules, and ions into the glomerulus as nephric filtrate. The essential ingredients are then reclaimed by the tubules, returning to the blood in the capillaries surrounding the tubules. The blood in these capillaries comes from the glomerulus (as in humans) and also from the renal portal veins which drain the posterior part of the fish's body.
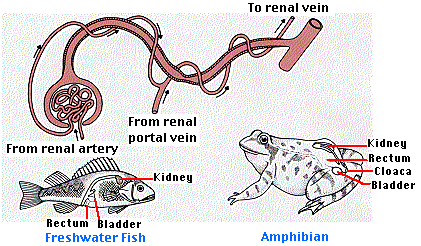
After solute reabsorption is complete, the urine is little more than water. Most of the nitrogenous wastes (including large amounts of ammonia, NH3) leave by diffusion out of the gills. So, the kidney is mostly a device for maintaining water balance in the animal, rather than an organ of excretion.
Amphibians
The amphibian kidney also functions chiefly as a device for excreting excess water. The permeable skin of the frog provides an easy route for the fresh water of its pond to enter by osmosis. But, as their name suggests, amphibians also spend time on land. Then the problem is to conserve water, not eliminate it.
The frog adjusts to the varying water content of its surroundings by adjusting the rate of filtration at the glomerulus. When blood flow through the glomerulus is restricted, a renal portal system is present to carry away materials reabsorbed through the tubules. The frog is also able to use its urinary bladder to aid water conservation. When in water, the frog's bladder quickly fills up with a hypotonic urine. On land, this water is reabsorbed into the blood helping to replace water lost through evaporation through the skin. The reabsorption is controlled by a hormone similar to mammalian ADH.
Lizards and Snakes
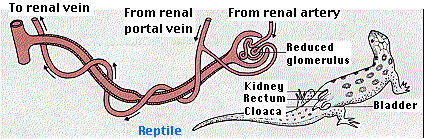
Many reptiles live in dry environments (e.g., rattlesnakes in the desert). Among the many adaptations to such environments is their ability to convert waste nitrogen compounds into uric acid. Uric acid is quite insoluble and so can be excreted using only a small amount of water. Thus we find that reptile glomeruli are quite small and, in fact, some reptiles have no glomeruli at all. Those with glomeruli filter just enough fluid to wash the uric acid, which is secreted by the tubules, into the cloaca. Most of this moisture is reabsorbed in the cloaca. Emptying the cloaca deposits feces (brown) and uric acid (a white paste). The cloaca is a chamber through which the feces and the gametes, as well as urine, pass on the way to the outside. The name comes from the Latin word for sewer. These water conservation mechanisms can allow the reptile to forgo drinking water. The water content of its food plus the water produced by cellular respiration is usually sufficient.
Birds
Bird kidneys function like those of reptiles (from which they are descended). Uric acid is also their chief nitrogenous waste. Most birds have a limited intake of fresh water. However, they need filter only enough to wash a slurry of uric acid into the cloaca where enough additional water is reclaimed to convert the uric acid into a semisolid paste. It is the whitish material that pigeons leave on statues.
Mammals
All mammals share our use of urea as their chief nitrogenous waste. Urea requires much more water to be excreted than does uric acid. Mammals produce large amounts of nephric filtrate but are able to reabsorb most of this in the tubules. But even so, humans lose several hundred ml each day in flushing urea out of the body. Some mammals have more efficient kidneys than ours. The kangaroo rat of the desert can produce a urine 17 times more concentrated that its blood. (The best we can do is 3-4 times as concentrated.) The efficiency of the kangaroo rat kidney enables it to survive without drinking water — simply depending on the water content of its food and that produced by cellular respiration.
We like to think of ourselves as highly advanced. Why don't we have kidneys as efficient as those of the reptiles and birds? It is the luck of our inheritance. The line of vertebrate evolution that produced the mammals split off before the evolution of the diapsids whose ability to convert nitrogenous wastes into uric acid was passed on to all their descendants, including the lizards, snakes, and birds.
Marine Fishes
Marine fishes face just the opposite problem from that of freshwater fishes. The salt content of sea water (~3%) is so hypertonic to that of their extracellular fluid that they are in continual danger of dehydration. The two major groups of marine fishes have solved this dilemma differently.
Cartilaginous Fishes (Chondrichthyes)
The cartilaginous fishes such as sharks, skates, and rays have developed high levels of urea in their blood. Shark's blood may contain 2.5% urea in contrast to the 0.01-0.03% in other vertebrates. This high level makes sharks blood isotonic to sea water, so the shark lives in osmotic balance with its environment and has a kidney that functions like ours with the exception that far more urea is reabsorbed in the shark's tubules than in ours.
Bony Fishes (Osteichthyes)
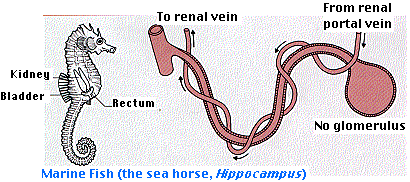
Marine bony fishes have solved the problem differently. They do lose water continuously but replace it by drinking sea water and then desalting it. The salt is returned to the sea by active transport at the gills. Living in constant danger of dehydration by the hypertonic sea, there is no reason to pump out large amounts of nephric filtrate at the glomerulus. The less water placed in the tubules, the less that has to be reabsorbed. So it is not surprising that many bony fishes have small glomeruli and some have no glomeruli at all. With a reduction in the filtration-reabsorption mechanism, the marine bony fishes rely more on tubular secretion for eliminating excess or waste solutes. Tubular secretion requires a good blood supply to the tubules. Lacking efficient glomeruli, the renal portal system must carry most of the burden.