
6.1.1.4: Food Chains and Food Webs
- Page ID
- 32156

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Trophic interactions in a community can be represented by diagrams called food chains and food webs. Before discussing these representations in detail, we must first review the basics of energy. Energy flows through a community as a result of trophic interactions.
Energy
Virtually every task performed by living organisms requires energy. In general, energy is defined as the ability to do work, or to create some kind of change. Energy exists in different forms. Examples include light energy, kinetic energy, heat energy, potential energy, and chemical energy.
When an object is in motion, there is energy associated with that object. Think of a wrecking ball. Even a slow-moving wrecking ball can do a great deal of damage to other objects. Energy associated with objects in motion is called kinetic energy. Heat energy is the energy of motion in matter (anything that takes up space and has mass) and is considered a type of kinetic energy. The warmer the substance, the faster its molecules are moving. The rapid movement of molecules in the air, a speeding bullet, and a walking person all have kinetic energy. Now what if that same motionless wrecking ball is lifted two stories above ground with a crane? If the suspended wrecking ball is not moving, is there energy associated with it? The answer is yes. The energy that was required to lift the wrecking ball did not disappear, but is now stored in the wrecking ball by virtue of its position and the force of gravity acting on it. This type of energy is called potential energy. If the ball were to fall, the potential energy would be transformed into kinetic energy until all of the potential energy was exhausted when the ball rested on the ground. Wrecking balls also swing like a pendulum; through the swing, there is a constant change of potential energy (highest at the top of the swing) to kinetic energy (highest at the bottom of the swing). Other examples of potential energy include the energy of water held behind a dam or a person about to skydive out of an airplane (figure \(\PageIndex{a}\)).
Potential energy is not only associated with the location of matter, but also with the structure of matter. Chemical energy is an example of potential energy that is stored in molecules. When molecules that are higher energy and less stable react to form products that are lower energy and more stable, this stored energy is released. Chemical energy is responsible for providing living cells with energy from food.
To appreciate the way energy flows into and out of biological systems, it is important to understand two of the physical laws that govern energy. The first law of thermodynamics states that the total amount of energy in the universe is constant and conserved. In other words, there has always been, and always will be, exactly the same amount of energy in the universe. Energy exists in many different forms. According to the first law of thermodynamics, energy may be transferred from place to place or transformed into different forms, but it cannot be created or destroyed. The transfers and transformations of energy take place around us all the time. Light bulbs transform electrical energy into light and heat energy. Gas stoves transform chemical energy from natural gas into heat energy. Plants perform one of the most biologically useful energy transformations on earth: that of converting the energy of sunlight to chemical energy stored within biological molecules, such as sugars (figure \(\PageIndex{b}\)).

The challenge for all living organisms is to obtain energy from their surroundings in forms that are usable to perform cellular work. A living cell’s primary tasks of obtaining, transforming, and using energy to do work may seem simple. However, the second law of thermodynamics explains why these tasks are harder than they appear. All energy transfers and transformations are never completely efficient. In every energy transfer, some amount of energy is lost in a form that is unusable. In most cases, this form is heat energy. For example, when a light bulb is turned on, some of the energy being converted from electrical energy into light energy is lost as heat energy. Likewise, some energy is lost as heat energy during the metabolic reactions that occur in organisms.
The concept of order and disorder relates to the second law of thermodynamics. The more energy that is lost by a system to its surroundings, the less ordered and more random the system is. Scientists refer to the measure of randomness or disorder within a system as entropy. High entropy means high disorder and low energy. Living things are highly ordered, requiring constant energy input to be maintained in a state of low entropy.
Energy Flow
Cells run on the chemical energy found mainly in carbohydrate molecules, and the majority of these molecules are produced by one process: photosynthesis. Through photosynthesis, certain organisms convert solar energy (sunlight) into chemical energy, which is then used to build carbohydrate molecules (figure \(\PageIndex{c}\)). The energy that is harnessed from photosynthesis enters the communities continuously and is transferred from one organism to another. Therefore, directly or indirectly, the process of photosynthesis provides most of the energy required by living things on Earth. See the Carbon Cycle and Photosynthesis in OpenStax Concepts of Biology for more details about photosynthesis.

Organisms that conduct photosynthesis (such as plants, algae, and some bacteria), and organisms that synthesize sugars through other means are called producers. Without these organisms, energy would not be available to other living organisms, and life would not be possible. Consumers, like animals, fungi, and various microorganisms depend on producers, either directly or indirectly. For example, a deer obtains energy by eating plants. A wolf eating a deer obtains energy that originally came from the plants eaten by that deer (figure \(\PageIndex{d}\)). Using this reasoning, all food eaten by humans can be traced back to producers that carry out photosynthesis (figure \(\PageIndex{e}\)).
Consumers can be classified based on whether they eat animal or plant material (figure \(\PageIndex{f}\)). Consumers that feed exclusively on animals are called carnivores. Lions, tigers, snakes, sharks, sea stars, spiders, and ladybugs are all carnivores. Herbivores are consumers that feed exclusively on plant material, and examples include deer, koalas, some bird species, crickets, and caterpillars. Herbivores can be further classified into frugivores (fruit-eaters), granivores (seed eaters), nectivores (nectar feeders), and folivores (leaf eaters). Consumers that eat both plant and animal material are considered omnivores. Humans, bears, chickens, cockroaches, and crayfish are examples of omnivores.
Dead producers and consumers are eaten by detritivores (which ingest dead tissues) and decomposers (which further break down these tissues into simple molecules by secreting digestive enzymes). Invertebrate animals, such as worms and millipedes, are examples of detritivores, whereas fungi and certain bacteria are examples decomposers.
Food Chains
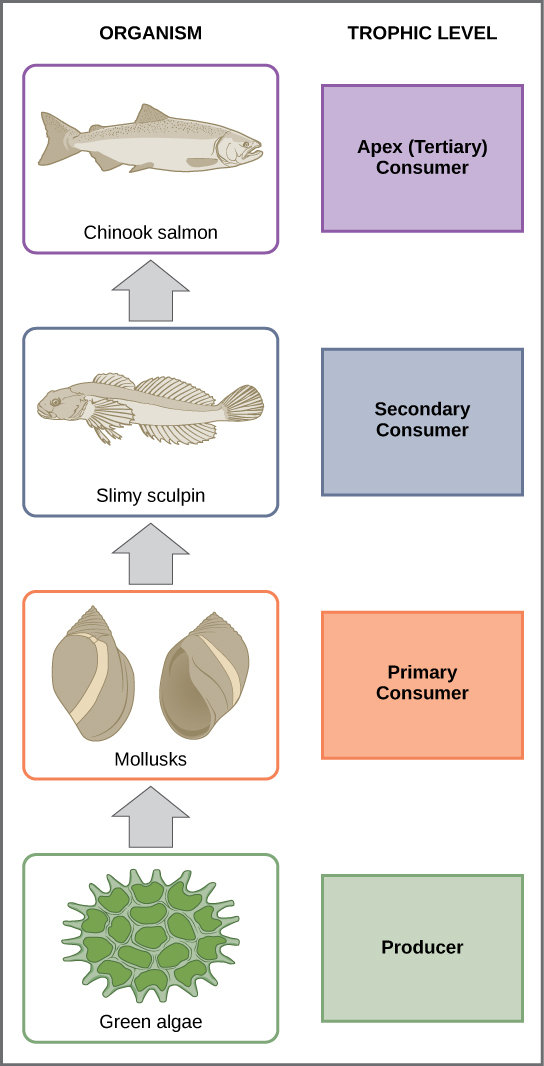
A food chain is a linear sequence of organisms through which nutrients and energy pass as one organism eats another. Each organism in a food chain occupies a specific trophic level (energy level), its position in the food chain. The first trophic level in the food chain is the producers. The primary consumers (the herbivores that eat producers) are the second trophic level. Next are higher-level consumers. Higher-level consumers include secondary consumers (third trophic level), which are usually carnivores that eat the primary consumers, and tertiary consumers (fourth trophic level), which are carnivores that eat other carnivores. In the Lake Ontario food chain, shown in figure \(\PageIndex{g}\), the Chinook salmon is the apex consumer at the top of this food chain. Some communities have additional trophic levels (quaternary consumers, fifth order consumers, etc.). Finally, detritivores and decomposers break down dead and decaying organisms from any trophic level. There is a single path through a food chain.
One major factor that limits the number of steps in a food chain is energy. Only about 10% of the energy in one trophic level is transferred to the next trophic level. This is because much energy is lost as heat during transfers between trophic levels or to decomposers due to the second law of thermodynamics. Thus, after four to six trophic energy transfers, the amount of energy remaining in the food chain may not be great enough to support viable populations at higher trophic levels (also see Community Productivity and Transfer Efficiency).
Certain environmental toxins can become more concentrated as they move up the food chain, with the highest concentrations occurring in the top consumers, a process called biomagnification. Essentially, a top consumer ingests all the toxins that had previously accumulated in the bodies of the organisms at the lower trophic levels. This explains why frequently eating certain fish, like tuna or swordfish, increases your exposure to mercury, a toxic heavy metal.
Food Webs
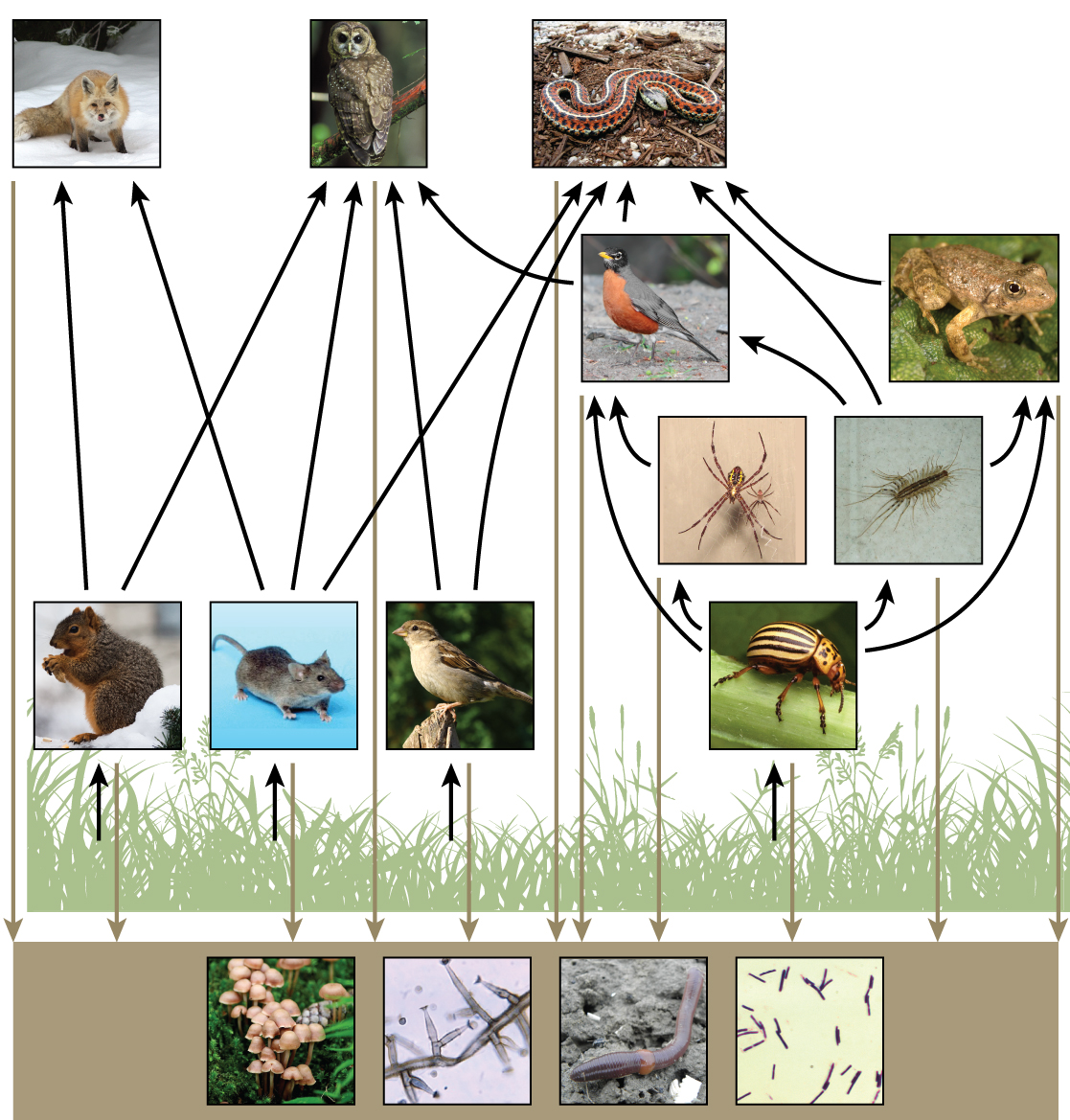
While food chains are simple and easy to analyze, there is a one problem when using food chains to describe most communities. Even when all organisms are grouped into appropriate trophic levels, some of these organisms can feed at more than one trophic level. In addition, species feed on and are eaten by more than one species. In other words, the linear model of trophic interactions, the food chain, is a hypothetical and overly simplistic representation of community structure. A holistic model—which includes all the interactions between different species and their complex interconnected relationships with each other and with the environment—is a more accurate and descriptive model. A food web is a concept that accounts for the multiple trophic interactions between each species (figure \(\PageIndex{h}\) and i).
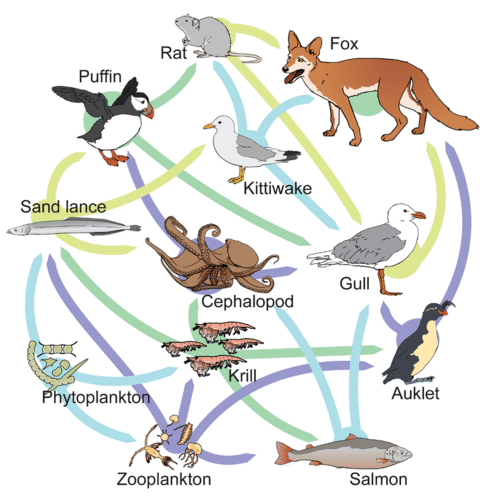
The trophic level of each species in a food web is not necessarily a whole number. In figure \(\PageIndex{i}\), phytoplankton are the primary producers (trophic level 1). Zooplankton only feed on phytoplankton, making them primary consumers (trophic level 2). Determining the trophic level of the other species is more complex. For example, krill eat both phytoplankton and zooplankton. If krill only ate phytoplankton they would primary consumers (trophic level 2). If they ate only zooplankton, they would be secondary consumers (trophic level 3). Since, krill consume both, their trophic level is 2.5.
Community Productivity and Transfer Efficiency
The rate at which photosynthetic producers incorporate energy from the sun is called gross primary productivity. In a cattail marsh, plants only trap 2.2% of the energy from the sun that reaches them. Three percent of the energy is reflected, and another 94.8% is used to heat and evaporate water within and surrounding the plant. However, not all of the energy incorporated by producers is available to the other organisms in the food web because producers must also grow and reproduce, which consumes energy. At least half of the 2.2% trapped by cattail marsh plants is used to meet the plants own energy needs.
Net primary productivity is the energy that remains in the producers after accounting for the metabolic needs of the producers and heat loss. The net productivity is then available to the primary consumers at the next trophic level. One way to measure net primary productivity is to collect and weigh the plant material produced on a m2 (about 10.7 ft2) of land over a given interval. One gram of plant material (e.g., stems and leaves), which is largely the carbohydrate cellulose, yields about 4.25 kcal of energy when burned. Net primary productivity can range from 500 kcal/m2/yr in the desert to 15,000 kcal/m2/yr in a tropical rain forest.
In an aquatic community in Silver Springs, Florida, the gross primary productivity (total energy accumulated by the primary producers) was 20,810 kcal/m2/yr (figure \(\PageIndex{j}\)). The net primary productivity (energy available to consumers) was only 7,632 kcal/m2/yr after accounting for energy lost as heat and energy require to meet the producer's metabolic needs.
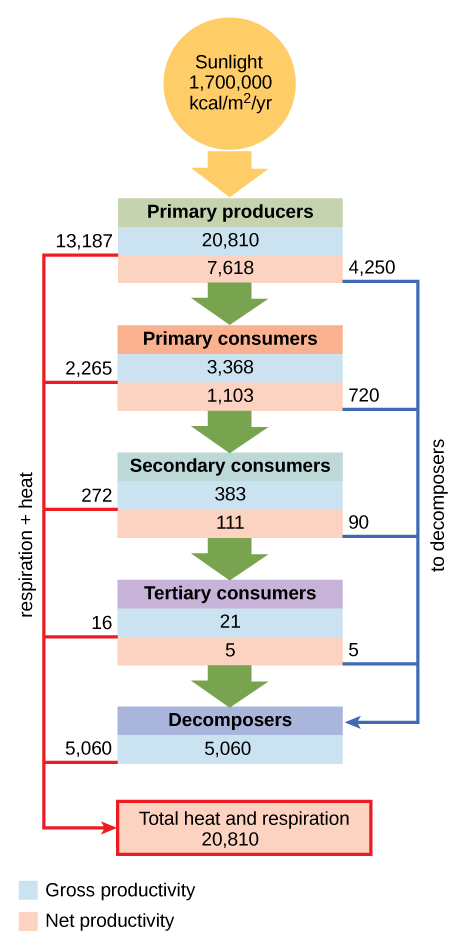
Only a fraction of the energy captured by one trophic level is assimilated into biomass, which makes it available to the next trophic level. Assimilation is the biomass of the present trophic level after accounting for the energy lost due to incomplete ingestion of food, energy used to conduct work by that trophic level, and energy lost as waste. Incomplete ingestion refers to the fact that some consumers eat only a part of their food. For example, when a lion kills an antelope, it will eat everything except the hide and bones. The lion is missing the energy-rich bone marrow inside the bone, so the lion does not make use of all the calories its prey could provide. In Silver Springs, only 1103 kcal/m2/yr from the 7618 kcal/m2/yr of energy available to primary consumers was assimilated into their biomass. (The trophic level transfer efficiency between the first two trophic levels was approximately 14.8 percent.)
An animal's source of heat influences its energy needs. Ectotherms, such as invertebrates, fish, amphibians, and reptiles, rely on external sources for body heat, and endotherms, such as birds and mammals, rely on internally generated heat. Generally, ectotherms require less of the energy to meet their metabolic needs and than endotherms do, and therefore, many endotherms have to eat more often than ectotherms.
The inefficiency of energy use by endotherms has broad implications for the world's food supply. It is widely accepted that the meat industry uses large amounts of crops to feed livestock, and because a low percentage of this is assimilated into biomass, much of the energy from animal feed is lost. For example, it costs about 1¢ to produce 1000 dietary calories (kcal) of corn or soybeans, but approximately $0.19 to produce a similar number of calories growing cattle for beef consumption. The same energy content of milk from cattle is also costly, at approximately $0.16 per 1000 kcal. Thus, there has been a growing movement worldwide to promote the consumption of non-meat and non-dairy foods so that less energy is wasted feeding animals for the meat industry.
Attribution
Modified by Melissa Ha from the following sources:
- Energy, Energy Enters Ecosystems Through Photosynthesis, and Energy Flow Through Ecosystems from Environmental Biology by Matthew R. Fisher (licensed under CC-BY)
- Ecology of Ecosystems, Energy Flow through Ecosystems, and Digestive Systems from General Biology/Biology 2e by OpenStax (licensed under CC-BY). Access for free at openstax.org.
- Ecosystem Productivity from Biology by John W. Kimball (licensed under CC-BY)