7.5: Origin of the Seed
- Page ID
- 18022

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
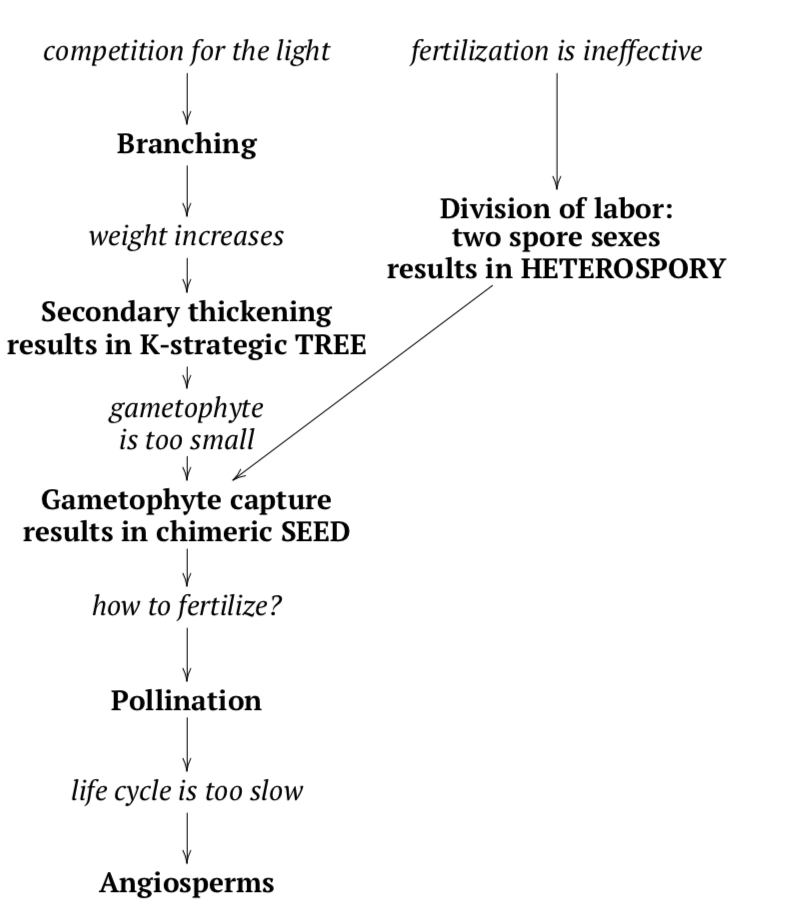
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)When plants developed the secondary growth, the almost unlimited perspectives opened for enlarging their body. However, these giants faced a new problem.
Big animals like elephants, lions, and whales tend to produce minimal number of offspring but increase the child care to ensure survival. This is called K-strategy, this is opposite to r-strategy usually smaller creatures which employs big numbers of offspring, and most of them will not survive (Figure \(\PageIndex{1}\)).

Analogously, bigger plants would need to do the same as K-strategic animals: make few daughter plants but defend and supply them with all needs until they mature. However, big secondary thickening spore plants were not capable of family planning; they still made billions of spores and then left them to fend for themselves. Naturally, only few from these billions would survive to become fertilized.
Spore reproduction is cheap and efficient but as birth control is not available, results are unpredictable. Even worse, these spore tree forests were not at all stable: in accidentally good conditions, many spores would survive and make sporophytes which start to grow simultaneously and then suppress each other and even die from over-population. But if the environmental conditions are bad, then none of the gametophytes will survive so there would be no new saplings to replace the old trees.
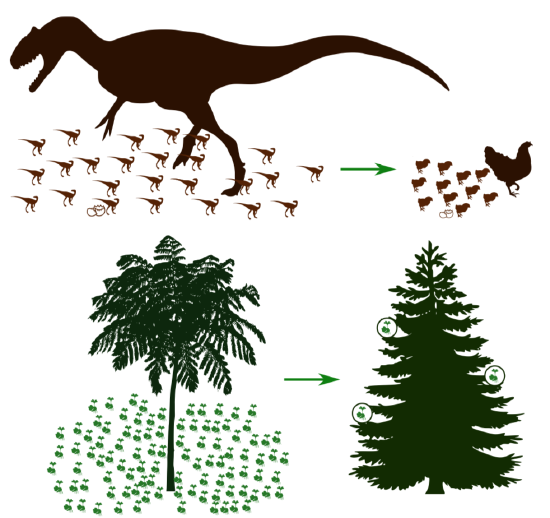
It is similar to the so-called “dinosaur problem”. This situation arose when giant Mesozoic reptiles also lost the control for their offspring: their egg size was limited due to physical restrictions, therefore, young dinosaurs were so much smaller than adults; then the only possible strategy was to leave them alone (Figure \(\PageIndex{2}\)). As a result, at the end of Mesozoic dinosaurs either decreased in size (became birds), or went extinct.
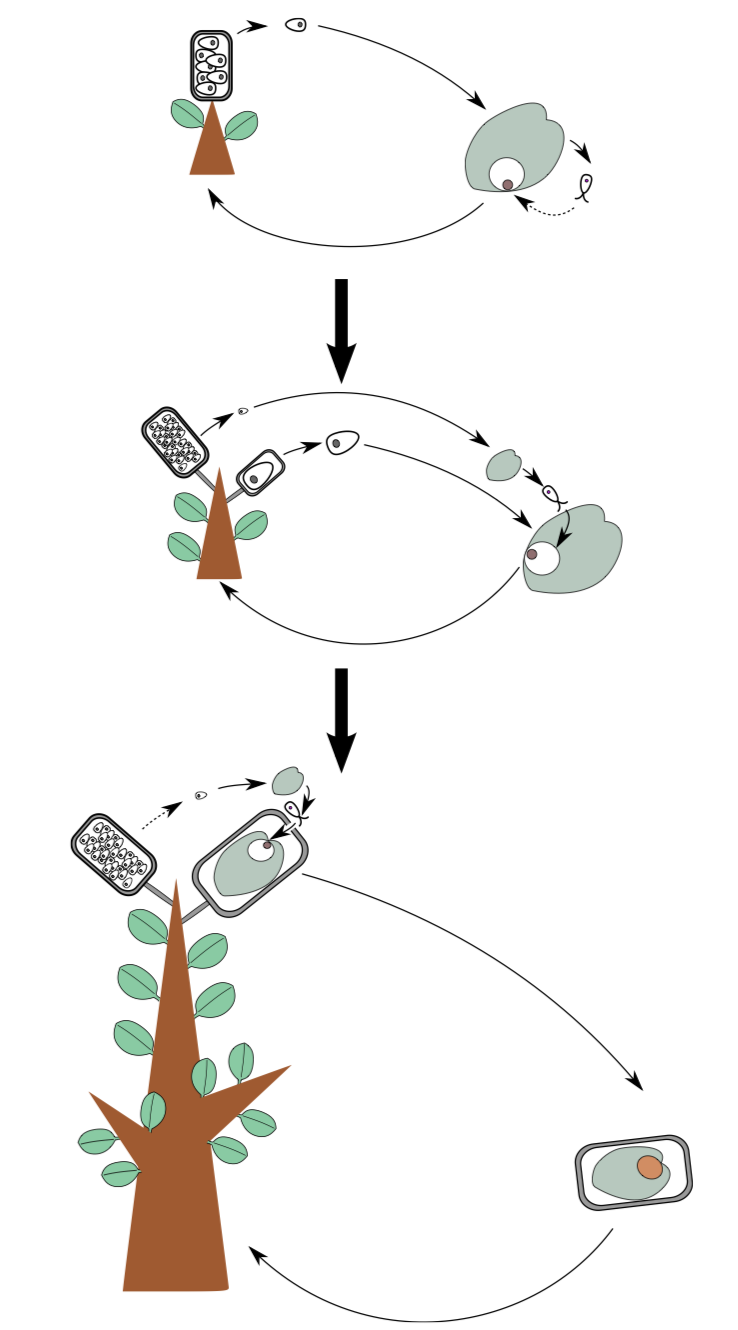
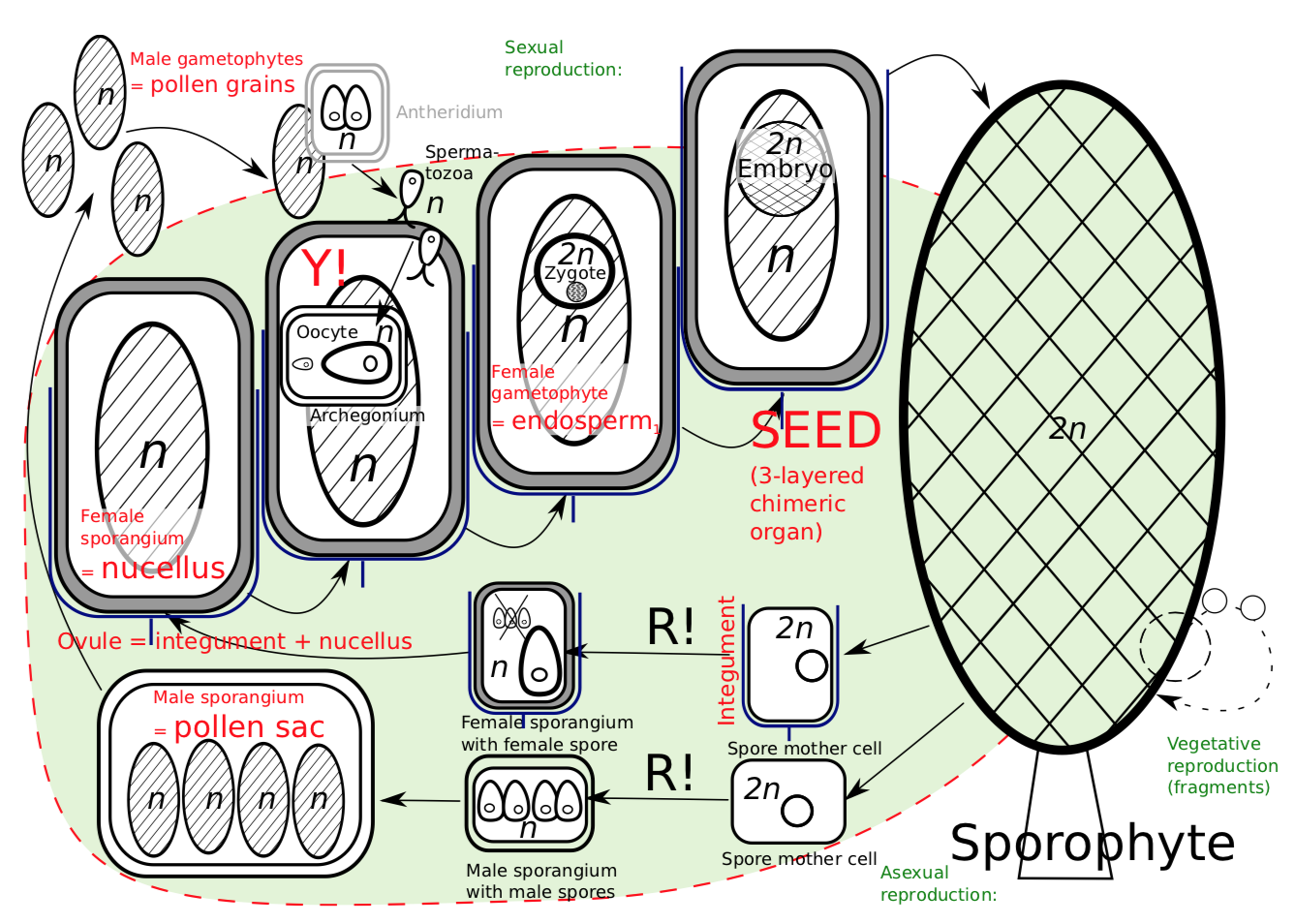
Plants, however, kept their size and survived. This is because they developed the seed (Figure \(\PageIndex{3}\)). A seed is the result of enforced control of the sporophyte over the gametophyte. The idea of a seed is to hide most of the heterosporous life cycle inside mother plant (Figure \(\PageIndex{3}\)). In seed plants, everything happens directly on the mother sporophyte: growing of gametophytes, syngamy, and growing of daughter sporophyte. Consequently, the female spore (megaspore) never leaves the sporangium. It germinates inside, waits for fertilization and then the zygote grows into and embryo, still inside the same sporangium.
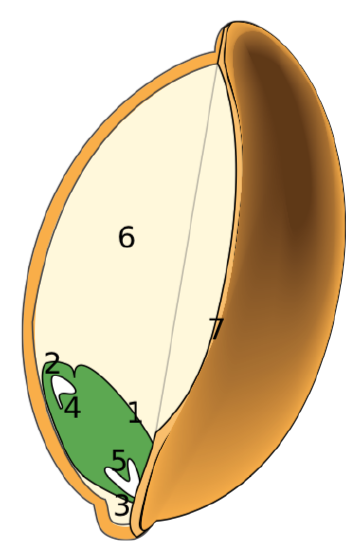
What will finally leave the mother plant is the whole female sporangium with gametophyte and embryo on it. This is the seedchimeric structure with mother (seed coat), daughter (embryo) and endosperm genotypes. It can be defined as chimeric structure with three genotypes: seed coat (mother plant megasporangium, \(2n\)), endosperm (female gametophyte, \(n\)), and daughter sporophyte (embryo, \(2n\)).
It should be noted here that flowering plants have endosperm of different origin; it is called endosperm\(_2\) and usually is triploid (\(3n\)) whereas female gametophyte endosperm is haploid (\(n\)) endosperm\(_1\)haploid nutrition tissue originated from female gametophyte. The other note is that apart from seed coat (which originates from integumentextra cover of megasporangium(s), megasporangium extra cover(s)), mother sporophyte also gives nucelluswall of megasporangium (wall of megasporangium) which sometimes is used as a feeding tissue for the embryo. This last tissue is called perispermnutrition tissue originated from nucellus (see).
One problem is still left. How will sperms reach female gametophyte and egg cell? The target is now high above the ground, on a branch of the giant tree. The only possible solution is pollination. Pollination is the distribution of the whole male gametophytes which are called pollen grains. Plants have no legs so they always need a third party in their sex, this is mostly wind or insects. A pollen grain is not a spore, mother sporophyte cares about male lineage too, and male spore grows into very small male gametophyte. It contains multiple haploid cells; some of which are sperms.
The lesser problem is: How would these sperms will swim to the egg cell? Some seed plants will excrete the drop of liquid from the top of the ovule (integument(s) + megasporangium), whereas the other, more advanced way is to grow a sperm delivery tool, the pollen tube (Figure \(\PageIndex{4}\)) made from one of the pollen grain cells. Fertilization with pollen tube is often called siphonogamy.
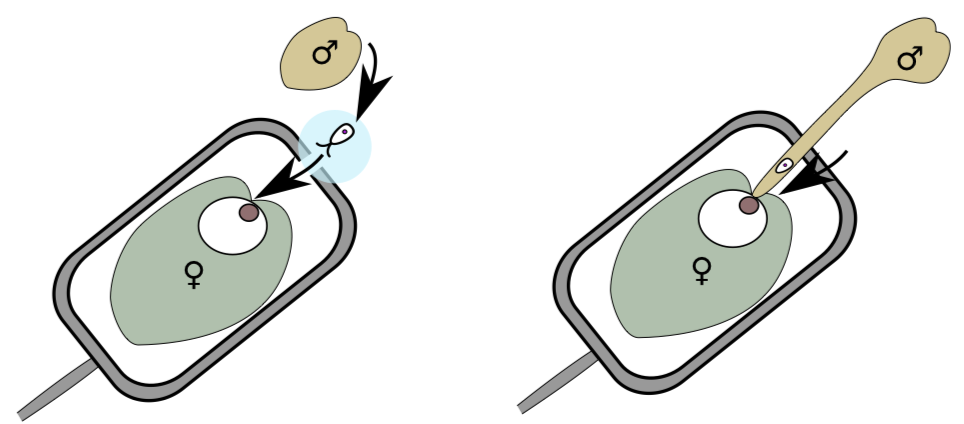
Consequently, seed plants with the pollen tube do not have flagella even on male gametes; these cells are spermatia: aflagellate, non-motile male gametes. (Below, we will continue to call all male gametes “sperms”). Pollen tube also allows only two male gametes per gametophyte: in living world, male gametes are usually competing for fertilization—this selects the best genotypes; whereas in higher seed plants, competition is between pollen tubes. Haploid pollen tube grows inside alien tissue of diploid sporophyte, so this growth is extremely slow in many seed plants. However, angiosperms made their pollen tubes grow fast.
With all these revolutionary adaptations, seed plants were first to colonize really dry places, and, in turn, allowed all other life to survive in arid climates.
The cycle of a seed plant (Figure \(\PageIndex{4}\)) begins with a sporophyte (\(2n\)) and has both the female and male organs where some cells undergo meiosis. Inside the ovule (which is the megasporangium with extra covers), female gametophyte (\(n\), future endosperm\(_1\)) produces the egg cells. Male gametophytes (pollen grains) ripen in the pollen sacseed plants: microsporangium which is the microsporangium. The pollen sac sends out the pollen grains which meet up with the ovule. The pollen grain then releases the sperms which fertilize the egg cell, and a zygote is formed. The zygote grows into embryo (which uses endosperm as a feeding tissue) and then into the sporophyte.
Several plant lineages met this “seed challenge”, there were seed lycophytes and also seed “horsetails”. However, seed ferns made it first and became ancestors of seed plants.
Seed Structure and Germination
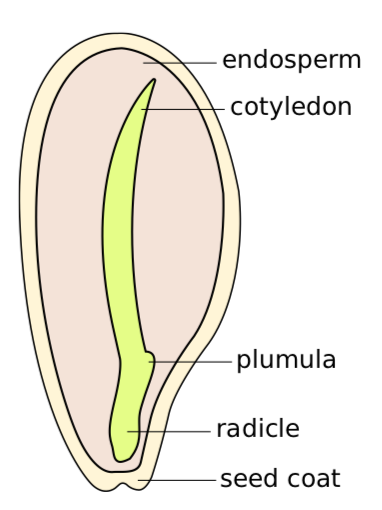
Seeds are diverse. For example, in an onion (Allium), a seed (Figure \(\PageIndex{6}\)) has endosperm, one cotyledonembryonic leaf (embryonic leaf), radicle (embryonic root), and the lateral embryonic bud (plumula).
Beans (Phaseolus) and other Leguminosae are examples of seeds without endosperm—actually, it was there, but growing embryo usually eats it out completely. These seeds have two large cotyledons. Grass (Gramineae) seeds contain several specific organs, namely coleoptile, coleorhiza, and scutellum. The scutellum is an enlarged cotyledon, coleoptile is the bud cover, and coleorhiza covers the embryonic root, radicle (Figure \(\PageIndex{7}\)). Onion and grasses are monocots with lateral embryonic bud. Other seed plants have a terminal embryonic bud and two or multiple cotyledons. Pine (Pinus) is an example of a plant that has multiple (five or more) cotyledons. Some plants like orchids (Orchidaceae) do not have developed embryo and even endosperm in seeds, their germination depends on a presence of symbiotic (mycorrhizal) fungus.
The first step in germination and starts with the uptake of water, also known as imbition. After imbition, enzymes are activated that start to break down starch into sugars consumed by embryo. The first indication that germination has begun is a swelling in the radicle. In onion and pea (Pisum), a structure that looks like a hook goes up through the soil and expose cotyledons and both hypocotyl and epicotyl (first internode). In beans, grasses, and palms, only epicotyl is exposed aboveground whereas cotyledons and hypocotyl remain underground.