
4.6.3: Mature Embryos and Seed Structure
- Page ID
- 32043

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)- Locate the major seed structures and identify the function of each.
- Compare eudicot and monocot seeds.
Eudicot Seeds
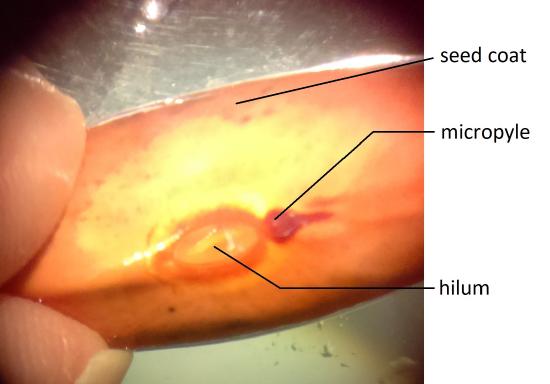
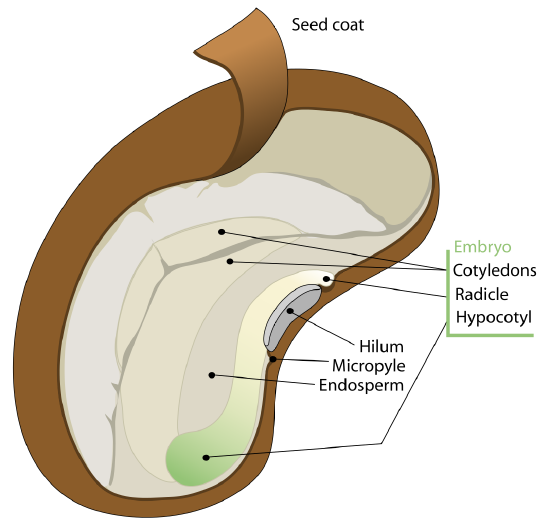
The seed is protected by a seed coat that is formed from the integument of the ovule (Figure \(\PageIndex{1}\)). The seed coat is further divided into an outer coat known as the testa and inner coat known as the tegmen. The hilum is a scar on the outside of the seed where it was attached to the endocarp (inner layer of the fruit wall). The micropyle is a small round structure next to the hilum where the pollen tube entered.
The embryonic axis (root-shoot axis) runs the length of the embryo. On end of the embryonic axis is the plumule, the young shoot apex, which includes the shoot apical meristem and developing leaves (leaf primordia). At the other end of the embryonic axis is the radicle (embryonic root). In some species, the radicle is not apparent in the embryo (in which case the distal end of the root is simply the root tip). The embryonic axis does not include the cotyledons. The portion of the embryo between the cotyledon attachment point and the radicle is known as the hypocotyl (hypocotyl means “below the cotyledons”). The portion of the embryonic axis between the cotyledon attachment and the shoot tip is the epicotyl (epicotyl means "above the cotyledons; Figures \(\PageIndex{2-3}\))). Some embryos lack a visible epicotyl because the cotyledons are attached to the embryonic axis at the shoot tip.
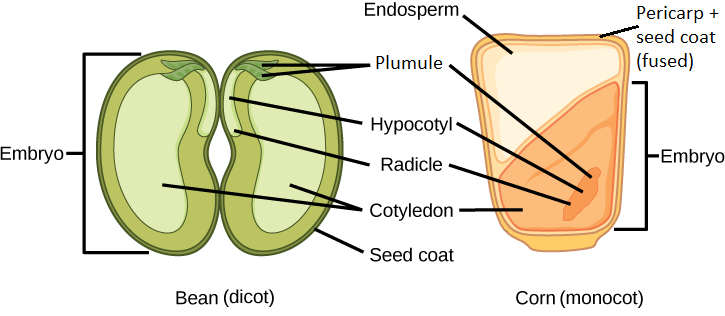
The two cotyledons in the eudicot seed are connected to the rest of the embryo via vascular tissue (xylem and phloem). In endospermic dicots, the food reserves are stored in the endosperm. During germination, the two cotyledons therefore act as absorptive organs to take up the enzymatically released food reserves. Tobacco (Nicotiana tabaccum), tomato (Solanum lycopersicum), and pepper (Capsicum annuum) are examples of endospermic dicots. In non-endospermic dicots, the triploid endosperm develops normally following double fertilization, but the endosperm food reserves are quickly remobilized and moved into the developing cotyledon for storage. The two halves of a peanut seed (Arachis hypogaea; Figure \(\PageIndex{4}\)) or a bean (Phaseolus; Figures \(\PageIndex{2-3}\)) and the split peas (Pisum sativum; Figure \(\PageIndex{5}\)) of split pea soup are individual cotyledons loaded with food reserves.


Because seeds have food reserves to fuel germination, they also are a nutritious food source for people. Studying the nutrient content of seed crops such as beans can be used to increase the nutritive value of the plant using biotechnology. Maria Elena Zavala (Figure \(\PageIndex{6}\)) is working to combat world hunger by manipulating plants to improve their nutritional qualities. For example, her research with beans is looking at how genetic engineering can be used to make the bean proteins more digestible and nutritious.

Monocot Seeds
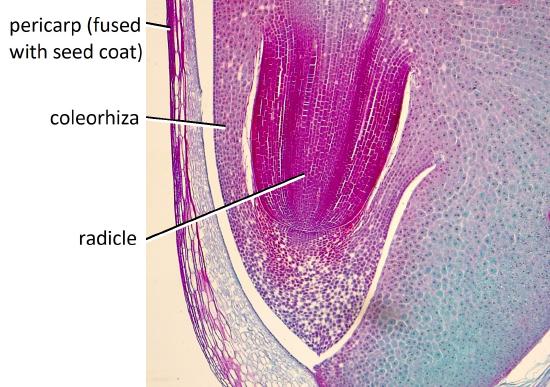
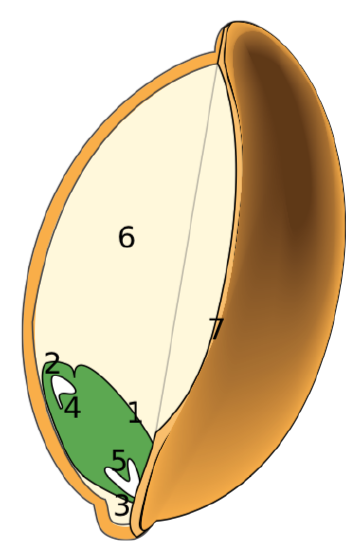
The seeds of the most complex monocot family, Poaceae (the grass family), which includes corn and wheat, are highly specialized. The testa and tegmen of the seed coat are fused. The fruit is a caryopsis (grain), a one-seeded fruit in which the fruit wall (pericarp) is fused to the seed coat. Thus, not only are the two layers of the seed coat fused, but the seed coat is fused to the pericarp.
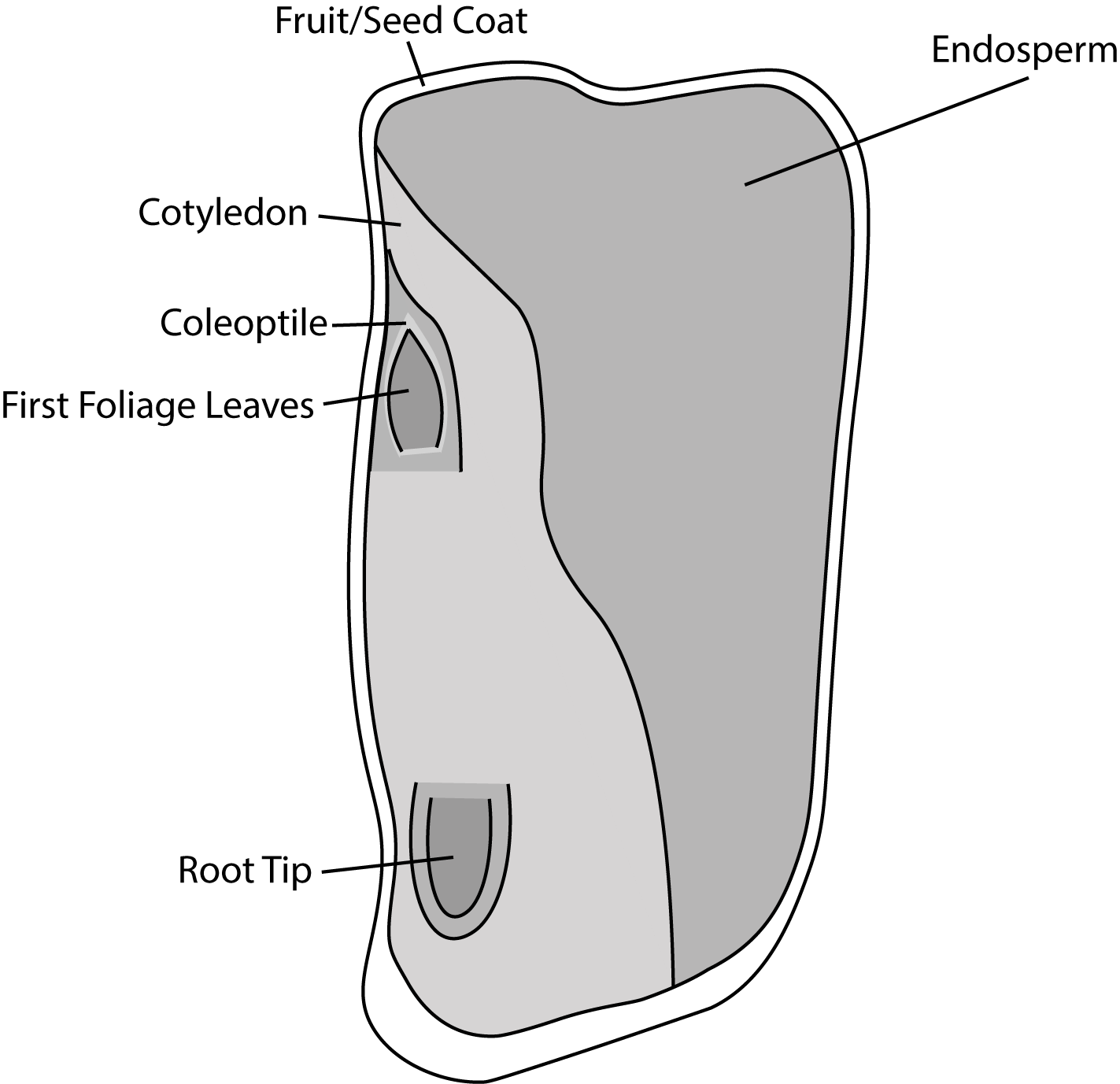
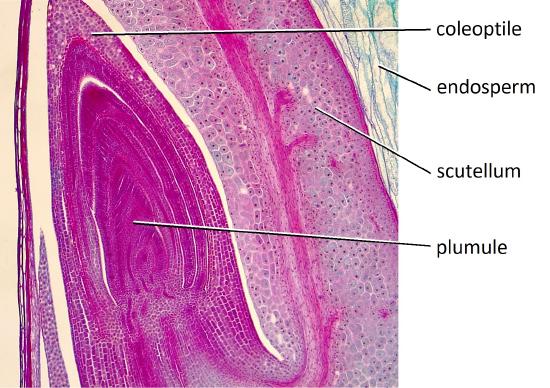
The single cotyledon is called a scutellum; the scutellum also has vascular connections to the rest of the embryo. The large inner layer of the endosperm that stores nutrients is called the starchy endosperm. The thin outer layer of the endosperm, which is a single layer of cells, is called the aleurone. Upon germination, enzymes are secreted by the aleurone. The enzymes degrade the stored carbohydrates, proteins and lipids, the products of which are absorbed by the scutellum and transported via a vasculature strand to the developing embryo. Therefore, the scutellum can be seen to be an absorptive organ, not a storage organ.
The root tip is protected by a sheath-like structure called the coleorhiza. Similarly, the coleoptile ensheaths the plumule at the shoot tip (Figure \(\PageIndex{7-10}\)).
Other Variations
Seeds are diverse. Pine (Pinus, a gymnosperm and thus neither a monocot nor eudicot) has multiple (five or more) cotyledons. Some plants like orchids (Orchidaceae, a monocot) do not have developed embryo and even endosperm in seeds; their germination depends on a presence of symbiotic (mycorrhizal) fungus.
Attributions
Curated and authored by Melissa Ha using the following sources:
- 32.2 Pollination and Fertilization from Biology 2e by OpenStax (licensed CC-BY). Access for free at openstax.org.
- 7.5 Origin of the Seed from Introduction to Botany by Alexey Shipunov (public domain)
- 16.3D Angiosperm Life Cycle from Biology by John W. Kimball (CC-BY)