
2.7.2: Pollination Syndromes
- Page ID
- 27726

Learning Objectives
- Explain pollination strategies with regard to genetic diversity.
- Explain what is meant by the term pollination syndrome.
- Use floral characteristics to predict a plants pollinator(s).
In angiosperms, pollination is defined as the placement or transfer of pollen from the anther to the stigma of the same flower or another flower. In gymnosperms, pollination involves pollen transfer from the male cone to the female cone. Upon transfer, the pollen germinates to form the pollen tube and the sperm for fertilizing the egg.
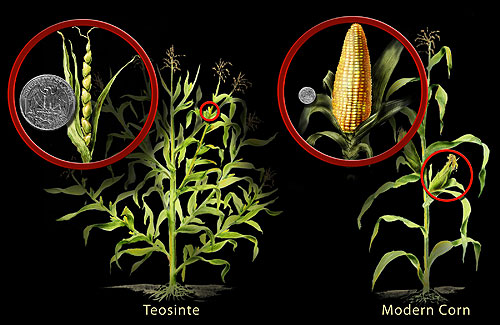
Pollination has been well studied since the time of Gregor Mendel. Mendel successfully carried out self- as well as cross-pollination in garden peas while studying how characteristics were passed on from one generation to the next. Today’s crops are a result of plant breeding, which employs artificial selection to produce the present-day cultivars (a term that refers to cultivated varieties). A case in point is today's corn, which is a result of years of breeding that started with its ancestor, a grass called teosinte (see Figure \(\PageIndex{1}\)). The teosinte that the ancient Mayans originally began cultivating had small seeds—vastly different from today’s relatively giant ears of corn and plump kernels. Interestingly, though these two plants appear to be entirely different, the genetic difference between them is miniscule.
.jpg?revision=1&size=bestfit&width=153&height=350)
.jpg?revision=1&size=bestfit&width=538&height=350)
Pollination and Genetic Diversity
Pollination takes two forms: self-pollination and cross-pollination. Self-pollination occurs when the pollen from the anther is deposited on the stigma of the same flower, or another flower on the same plant. Self-pollination occurs in flowers where the stamen and carpel mature at the same time, and are positioned so that the pollen can land on the flower’s stigma. This method of pollination does not require an investment from the plant to provide nectar and pollen as food for pollinators, but results in lower genetic diversity within the population. Self-pollination often exists like a “plan B”, in case cross-pollination is, for some reason, impossible. Sometimes, self-pollinated flowers even do not open; these flowers are called cleistogamous.
Cross-pollination is the transfer of pollen from the anther of one flower to the stigma of another flower on a different individuals of the same species. This method relies on a pollen vector but results in higher genetic diversity.
Living species have adapted to ensure survival of their progeny; those that fail become extinct. Genetic diversity is therefore required so that in changing environmental or stress conditions, some of the progeny can survive. Self-pollination leads to the production of plants with less genetic diversity, since genetic material from the same plant is used to form gametes, and eventually, the zygote. In contrast, cross-pollination—or out-crossing—leads to greater genetic diversity because the microgametophyte and megagametophyte are derived from different plants.
Because cross-pollination allows for more genetic diversity, plants have developed many ways to avoid self-pollination. In some species, the pollen and the ovary mature at different times. These flowers make self-pollination nearly impossible. By the time pollen matures and has been shed, the stigma of this flower is mature and can only be pollinated by pollen from another flower. Some flowers have developed physical features that prevent self-pollination. The primrose is one such flower. Primroses have evolved two flower types with differences in anther and stigma length: the pin-eyed flower has anthers positioned at the pollen tube’s halfway point, and the thrum-eyed flower’s stigma is likewise located at the halfway point. Insects easily cross-pollinate while seeking the nectar at the bottom of the pollen tube. This phenomenon is also known as heterostyly. Many plants, such as cucumber, have male and female flowers located on different parts of the plant, thus making self-pollination difficult. In yet other species, the male and female flowers are borne on different plants (dioecious). All of these are barriers to self-pollination; therefore, the plants depend on pollinators to transfer pollen. The majority of pollinators are biotic agents such as insects (like bees, flies, and butterflies), bats, birds, and other animals. Other plant species are pollinated by abiotic agents, such as wind and water.
In summary, self-incompatibility is a mechanism that prevents self-fertilization in many flowering plant species to increase genetic diversity at the population level. The working of this self-incompatibility mechanism has important consequences for plant breeders because it inhibits the production of inbred and hybrid plants.
Incompatibility Genes in Flowers
In recent decades, incompatibility genes—which prevent pollen from germinating or growing into the stigma of a flower—have been discovered in many angiosperm species. If plants do not have compatible genes, the pollen tube stops growing. Self-incompatibility is controlled by the S (sterility) locus. Pollen tubes have to grow through the tissue of the stigma and style before they can enter the ovule. The carpel is selective in the type of pollen it allows to grow inside. The interaction is primarily between the pollen and the stigma epidermal cells. In some plants, like cabbage, the pollen is rejected at the surface of the stigma, and the unwanted pollen does not germinate. In other plants, pollen tube germination is arrested after growing one-third the length of the style, leading to pollen tube death. Pollen tube death is due either to apoptosis (programmed cell death) or to degradation of pollen tube RNA. The degradation results from the activity of a ribonuclease encoded by the S locus. The ribonuclease is secreted from the cells of the style in the extracellular matrix, which lies alongside the growing pollen tube.
Pollination Syndromes
Over time, angiosperms evolved different flower morphologies, smells, and colors that corresponded to their particular pollen vector. These sets of characteristics, called pollination syndromes, allow scientists to predict the pollinators for different plants. For example, cup-shaped flowers are usually pollinated with massive animals like beetles and even bats. Funnel-shaped flowers as well as labiate flowers (with lips), are adapted to flies and bees. Flowers with long spurs attract butterflies and birds (like hummingbirds or sugarbirds). Likewise, many of the organismal vectors evolved features or behaviors to specialize in pollination of particular plants. This tandem evolution toward increasing specificity is called coevolution. Interestingly, flower phenotypes can also correlate to other environmental factors (e.g. Peach et al. 2020). For more images of pollination syndromes, see Chapter 14.1 Pollination Syndromes in the Photographic Atlas.
Pollination by Deception
Orchids are highly valued flowers, with many rare varieties (Figure \(\PageIndex{2}\)). They grow in a range of specific habitats, mainly in the tropics of Asia, South America, and Central America. At least 25,000 species of orchids have been identified.

Flowers often attract pollinators with food rewards, in the form of nectar. However, some species of orchid are an exception to this standard: they have evolved different ways to attract the desired pollinators. They use a method known as food deception, in which bright colors and perfumes are offered, but no food. Anacamptis morio, commonly known as the green-winged orchid, bears bright purple flowers and emits a strong scent. The bumblebee, its main pollinator, is attracted to the flower because of the strong scent—which usually indicates food for a bee—and in the process, picks up the pollen to be transported to another flower.
Other orchids use sexual deception. Chiloglottis trapeziformis emits a compound that smells the same as the pheromone emitted by a female wasp to attract male wasps. The male wasp is attracted to the scent, lands on the orchid flower, and in the process, transfers pollen. Some orchids, like the Australian hammer orchid, use scent as well as visual trickery in yet another sexual deception strategy to attract wasps. The flower of this orchid mimics the appearance of a female wasp and emits a pheromone. The male wasp tries to mate with what appears to be a female wasp, and in the process, picks up pollen, which it then transfers to the next counterfeit mate.
Bees
Bees are perhaps the most important pollinator of many garden plants and most commercial fruit trees (Figure \(\PageIndex{3}\)). Most bees that people notice are the social bees, bumblebees and honeybees, though there are thousands of species of solitary bees that have essential pollination roles. Bees have branched hairs covering their bodies, making them excellent pollinators as more pollen grains are likely to be caught upon their external surfaces and transferred onto other plants.
Bees collect energy-rich pollen and nectar for their survival and energy needs, so flowers that have coevolved with bees contain both nectar and pollen. Since bees cannot see the color red, bee-pollinated flowers usually have shades of blue, yellow, violet or other colors. They visit flowers that are open during the day, are brightly colored, have a strong aroma or scent, with a landing area of some kind, typically with the presence of a nectar guide. A nectar guide can include regions on the flower with pigments that are visible only to bees, and not to humans (such as UV); it helps to guide bees to the center of the flower, thus making the pollination process more efficient. The pollen sticks to the bees’ fuzzy hair, and when the bee visits another flower, some of the pollen is transferred to the second flower.


Recently, populations of honeybees (an introduced European species) have been in decline due to a variety of factors, including pesticides (like neonicotinoids) and parasitic mites. Because solitary bees are not operated by commercial industry, impacts on these populations have been less well-documented. However, declines in bee populations will have extremely important impacts on our food security, as bees are used to pollinate many food crops.
Wasps are also important insect pollinators, and pollinate many species of figs.
Flies
Many fly species are important pollinators. These flies often look and behave much like bees, performing important roles in pollination. However, since these flowers would generally be classified under a bee pollination syndrome, we will instead discuss a second category of fly for the fly pollination syndrome. These insects (flies and sometimes carrion beetles) are typically not looking for a flower, but for a corpse or some dung to feed on or lay eggs in. Some flowers have evolved to trick flies into pollination by emitting a decaying smell or an odor of rotting flesh. These flowers, which produce nectar, usually have dull colors, such as brown or purple. They often feature some kind of trap-like feature so when a fly visits, it has to bounce around inside the flower a bit before leaving (this increases the chance of pollen deposition, both on the flower and on the fly). Examples include the corpse flowers (Amorphophallus and Rafflesia) and dragon arum (Dracunculus).

Butterflies and Moths
Butterflies, such as the monarch, pollinate many garden flowers and wildflowers, which usually occur in clusters. These flowers are brightly colored, have a strong fragrance, are open during the day, and have nectar guides to make access to nectar easier. The pollen is picked up and carried on the butterfly’s limbs.
Moths, on the other hand, pollinate flowers during the late afternoon and night. The flowers pollinated by moths are pale or white and are flat, enabling the moths to land. One well-studied example of a moth-pollinated plant is the yucca plant, which is pollinated by the yucca moth. The shape of the flower and moth have adapted in such a way as to allow successful pollination. The moth deposits pollen on the sticky stigma for fertilization to occur later. The female moth also deposits eggs into the ovary. As the eggs develop into larvae, they obtain food from the flower and developing seeds. Thus, both the insect and flower benefit from each other in this symbiotic relationship. The corn earworm moth and Gaura plant have a similar relationship (Figure \(\PageIndex{5}\)).

Bats
In the tropics and deserts, bats are often the pollinators of nocturnal flowers such as agave, guava, and morning glory. The flowers are usually large and white or pale-colored; thus, they can be distinguished from the dark surroundings at night. The flowers have a strong, fruity, or musky fragrance and produce large amounts of nectar. They are naturally large and wide-mouthed to accommodate the head of the bat (Figure \(\PageIndex{6}\)). As the bats seek the nectar, their faces and heads become covered with pollen, which is then transferred to the next flower.

Birds
Many species of small birds, such as the hummingbird (Figure \(\PageIndex{7}\)) and sun birds, are pollinators for plants such as orchids and other wildflowers. Flowers visited by birds are usually sturdy and are oriented in such a way as to allow the birds to stay near the flower without getting their wings entangled in the nearby flowers, such as dangling (pendant). The flower typically has a curved, tubular shape, which allows access for the bird’s beak. Brightly colored, odorless flowers that are open during the day are pollinated by birds. As a bird seeks energy-rich nectar, pollen is deposited on the bird’s head and neck and is then transferred to the next flower it visits. Botanists have been known to determine the range of extinct plants by collecting and identifying pollen from 200-year-old bird specimens from the same site.

Wind
Most species of conifers, and many angiosperms, such as grasses, maples and oaks, are pollinated by wind. Pine cones are brown and unscented, while the flowers of wind-pollinated angiosperm species are usually green, small, may have small or no petals, and produce large amounts of pollen. Unlike the typical insect-pollinated flowers, flowers adapted to pollination by wind do not produce nectar or scent. In wind-pollinated angiosperm species, the anthers are often large and hang out of the flower, and, as the wind blows, the lightweight pollen is carried with it (Figure \(\PageIndex{8}\)). The flowers usually emerge early in the spring, before the leaves, so that the leaves do not block the movement of the wind. The pollen is deposited on the exposed feathery stigma of the flower (Figure \(\PageIndex{9}\)).


Water
Some plants, such as Australian sea grass and pond weeds, are pollinated by water. The pollen floats on water, and when it comes into contact with the flower, it is deposited inside the flower.
Summary Table of Pollination Syndromes
Table \(\PageIndex{1}\) summarizes the pollination syndromes described in the categories above.
| Pollination Syndrome | Color | Structure | Scent | Nectar or Pollen |
|---|---|---|---|---|
| Wind | Dull, perianth often absent or reduced | Large feathery stigmas, large anthers | None | No nectar, large amounts of pollen |
| Birds | Reds and pinks | Often tubular or cupped | None | Lots of hidden nectar, moderate pollen |
| Bees | Purples, blues, yellows, white, UV | Flat and shallow or tubular, with landing area | Sweet, fresh, mild | Pollen often sticky and scented, nectar usually present |
| Bats | White, dull green, or purple | Often bowl-shaped or pendant, anthers protruding | Musty or fruity, strong, emitted at night | Lots of hidden nectar |
| Moths | White, pale pink or purple | Often tubular or cupped, no landing pad | Strong and sweet, emitted at night | Lots of hidden nectar, limited pollen |
| Butterflies | Bright colors | Tubular, with wide landing pad | Faint, fresh | Lots of hidden nectar, limited pollen |
| Flies | Dark red, purple, brown | Shallow, funnel, or trap-like | Putrid, rotting | No nectar, moderate pollen |
Attributions
Curated and authored by Maria Morrow using the following sources:
- 32.2 Pollination and Fertilization from Biology 2e by OpenStax (licensed CC-BY). Access for free at openstax.org.
- 8.2 The Flower and the Fruit from Introduction to Botany by Alexey Shipunov (public domain)