
2.5: Gene and Operon
- Page ID
- 18130

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Genes
The entire nucleic acid sequence that is necessary for the synthesis of a functional polypeptide or RNA molecule. Thus, a gene contains additional sequence information beyond that which codes for the amino acids in a protein or the nucleotides in an RNA molecule. The gene also contains the DNA necessary to get a particular transcript made.
Transcription control regions can be remote to the coding region (on the order of Kb's or 10's of Kb's away).
Most prokaryotic genes lack introns (intervening DNA sequence). In prokaryotes, genes which encode proteins with relationships in a metabolic pathway form Operons - which produce polycistronic mRNA's.
Definitions: Operon and Promoter
- An operon is in bacterial DNA, a cluster of contiguous genes transcribed from one promoter that gives rise to a polycistronic mRNA.
- A Promoter is a DNA sequence to which RNA polymerase binds prior to initiation of transcription - usually found just upstream of the transcription start site of a gene
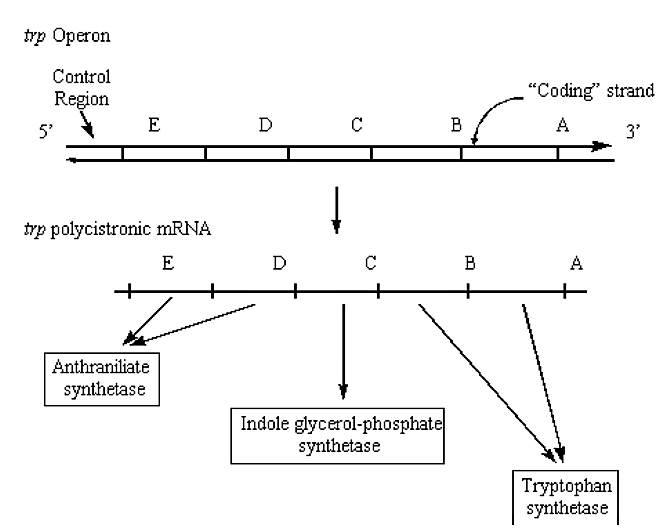
e.g. Trp Operon - involved in the biosynthesis of the amino acid tryptophan:
.png?revision=1&size=bestfit&width=486&height=442)
Figure 2.5.1: Chemical pathway of trp operon
.png?revision=1&size=bestfit&width=463&height=369)
Figure 2.5.2: Trp operon in DNA/RNA
A consequence of the arrangement of bacteria genes into operons is that the level of mRNA for each of the genes in the operon is exactly the same.
Ribosomes transcribe from the start of each gene, not only from the first gene.
Another consequence of the arrangement of bacteria genes into operons is that an upstream mutation (i.e. possibly inhibiting transcription) can prevent "downstream" genes from being transcribed and expressed. Most eukaryotic transcription units produce monocistronic mRNA's, (i.e. they encode only one protein). There is a fundamental difference in the translation processes of prokaryotes and eukaryotes:
- In prokaryotes ribosomes can bind at specific recognition sequences anywhere within the mRNA (called ribosome binding sites, or "Shine-Dalgarno" sites).
- In eukaryotes, ribosomes bind via the interaction with specifically modified 5' region (so called 5' cap site) of mRNA molecules.
- Most eukaryotic mRNA's are therefore monocistronic.
- Mutations in simple eukaryotic transcription units affect only one protein.
Complex Eukaryotic Transcription Units
The primary RNA transcript encoded by complex transcription units can be spliced in more than one way. Because of the different processing possibilities, the exons (coding regions) in a single complex transcription unit can be linked in alternative ways, to yield different mRNAs and different proteins.
.png?revision=1&size=bestfit&width=588&height=414)
Figure 2.5.3: Complex transcription units
Transcriptional regulation
Successful survival requires adaptability and economy:
- The ability to switch from metabolizing one substrate to another as environmental resources change
- It would be an energetic waste to produce enzymes for a metabolic pathway which is not needed.
Induction versus Repression of Enzyme Synthesis
In E. coli certain enzymes are produced only when the cells are grown on certain substrates. This effect is called enzyme induction. For example, when cells are grown in the absence of a type of sugar known as a b galactoside (e.g. lactose) the cells contain very few molecules (~5 per cell) of the enzyme b-galactosidase (which cleaves lactose into glucose and galactose).
- There is no need for this enzyme in the absence of lactose.
- If lactose is added to E. coli, in a very short amount of time there are approximately 5000 molecules of b-galactosidase per cell (approximately ~1,000 fold induction).
- If lactose is removed from the media synthesis of b-galactosidase stops.
A similar but opposite situation occurs in regard to the synthesis of tryptophan (the biosynthetic enzymes are contained in the trp operon). In this case production of the enzymes for tryptophan biosynthesis are rapidly shut down if tryptophan is present, in a process called repression. Repression is a transcriptional regulatory mechanism for commonly required gene products. Induction is a transcriptional regulatory mechanism for gene products which may be required under unusual or infrequent situations.