
7.1: Monosaccharides and Disaccharides
- Page ID
- 102271


\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)
(Learning goals written by Claude, Sonnet 4.6, Anthropic)
Monosaccharide Structure and Stereochemistry
- Define carbohydrates as polyhydroxy aldehydes or ketones, distinguish aldoses from ketoses and classify sugars by carbon number (trioses through hexoses), explain the D/L designation based on the configuration of the asymmetric center most remote from the carbonyl relative to D-glyceraldehyde, and interconvert Fischer projections, wedge-dash representations, Haworth projections, and chair conformations of common hexoses — applying the rules that substituents pointing right in Fischer projections point down in Haworth projections, that α-anomers project down (αnts down) and β-anomers project up (βutterflies up) in Haworth projections, and that β-D-glucopyranose is uniquely stable because all bulky substituents occupy equatorial positions in the chair conformation.
- Distinguish and provide examples of the four types of sugar configurational isomers — enantiomers (mirror images, e.g., D-Glc vs. L-Glc), diastereomers (non-mirror-image stereoisomers), epimers (diastereomers differing at a single stereocenter, e.g., glucose and galactose differ at C4), and anomers (epimers at the anomeric carbon formed during ring closure, e.g., α-D-glucopyranose vs. β-D-glucopyranose) — and explain why anomers interconvert in solution (mutarotation) through the open-chain aldehyde intermediate, while enantiomers, diastereomers, and non-anomeric epimers require covalent bond breaking to interconvert.
- Describe the mechanism of pyranose and furanose ring formation as an intramolecular nucleophilic attack of a hydroxyl group on the aldehyde or ketone carbonyl carbon, forming a hemiacetal with the anomeric hydroxyl pointing either α (down) or β (up) — and explain why glucose exists predominantly as β-D-glucopyranose in solution (75% of the equilibrium mixture) and exclusively as pyranose in polysaccharides, while fructose and ribose prefer the furanose form in nucleotides and polynucleotides.
Monosaccharide Derivatives and Glycosidic Bond Chemistry
- Identify the major chemical modifications found in monosaccharide derivatives — including oxidation of the aldehyde to gluconate (C1) or the terminal alcohol to glucuronate (C6), phosphorylation at C1 or C6 by ATP-dependent kinases, amination to glucosamine and galactosamine, N-acetylation to GlcNAc and GalNAc, and condensation with lactate (muramic acid, bacterial cell walls) or pyruvate (neuraminic/sialic acids, mammalian cell surfaces) — and explain the biosynthetic origin and biological significance of each derivative.
- Explain the mechanism of glycosidic bond formation as acid-catalyzed acetal formation — in which protonation of the anomeric OH generates water as a leaving group, enabling nucleophilic attack by a second hydroxyl (from another monosaccharide) to form the acetal linkage — and apply this to describe the structural diversity of disaccharides (maltose: Glc(α1→4)Glc; cellobiose: Glc(β1→4)Glc; lactose: Gal(β1→4)Glc; sucrose: Glc(α1→2β)Fru) in terms of the identity of the sugars, the anomeric configuration, and the carbon positions involved in the linkage.
- Distinguish reducing from nonreducing sugars by explaining that any mono- or disaccharide with a free hemiacetal (anomeric OH available to open to an aldehyde) is a reducing sugar capable of reducing Cu²⁺ to Cu²O in Benedict's or Fehling's reagent — whereas sucrose, in which both anomeric carbons are involved in the glycosidic link (a full acetal), cannot open to an aldehyde and is therefore nonreducing — and connect this to the clinical use of urine glucose tests and to alpha-gal syndrome, where the disaccharide Gal(α1→3)Gal in tick saliva triggers IgE-mediated hypersensitivity reactions that cross-react with red meat glycans.
Introduction
Carbohydrate or glycan biochemistry is very complex and challenging owing to the stereochemical complexity of simple sugars, the large number of positions on the sugars used to form linkages between other sugars to create polymers, the large number of chemical modifications to base sugars, and the lack of a genetic template to instruct glycan polymer formation. No wonder our understanding of complex glycans has developed more slowly than that of chemically simpler polymers like nucleic acids and proteins.
In addition, the terminology used to describe them varies. We use these general descriptions of them:
Sugar: usually refers to low molecular weight carbohydrates like glucose, lactose, and sucrose, but it can also refer broadly to any carbohydrate.
Carbohydrate: a general term that applies to simple sugars to complex sugar polymers like glycogen, starch, and cellulose. The name derives from the formula for simple sugars like glucose (C6H12O6), which can be written as C6(H2O)6 - a carbo (C) - hydrate (H2O).
Glycan: a general term for molecules containing simple sugars and sugar derivatives linked in a polymer, either standalone molecules or attached to other molecules like proteins.
Monosaccharides Structures
The above definition of sugar needs some further nuance. From a chemical perspective, sugars are polyhydroxyl aldehydes or ketones. The simplest sugars contain at least three carbon atoms, and the most common are the aldo- and keto-trioses, tetroses, pentoses, and hexoses. The 3C sugars are glyceraldehyde and dihydroxyacetone, as shown in Figure \(\PageIndex{1}\).

Figure \(\PageIndex{1}\): Three-carbon sugars
Glucose, an aldohexose, is a central sugar in metabolism. It and other 5C and 6C sugars can cyclize through intramolecular nucleophilic attack of one of the free hydroxyl groups on the carbonyl carbon of the aldehyde or ketone. Such intramolecular reactions occur if stable 5- or 6-member rings can form. The resulting rings are labeled furanose (5-member) or pyranose (6-member) based on their similarity to furan and pyran. On nucleophilic attack to form the ring, the carbonyl O becomes an OH that points either below (α anomer) or above (β anomer) the ring.
Figure \(\PageIndex{2}\) shows different representations of the linear and cyclic forms of the sugars D-glucose, D-ribose, and D-fructose

Monosaccharides in solution exist as equilibrium mixtures of the straight and cyclic forms. In solution, glucose (Glc) is mainly in the pyranose form, fructose is 67% pyranose and 33% furanose, and ribose is 75% furanose and 25% pyranose. However, in polysaccharides, Glc is exclusively pyranose, and fructose and ribose are furanoses.
Sugars can be drawn in straight-chain form as Fischer projections or perspective structural formulas.
In the Fischer projection, the vertical bonds point down into the plane of the paper. That's easy to visualize for 3C sugars, but more complicated for larger ones. For those, draw a wedge-and-dash line drawing of the molecule. When determining the orientation of the OHs on each C, orient the wedge and dash drawing in your mind so that the C atoms adjacent to the one of interest are pointing down. Sighting towards the carbonyl C, if the OH is pointing to the right in the Fischer project, it should be pointing to the right in the wedge and dash drawing, as shown below for D-threose and D-glucose. Figure \(\PageIndex{3}\) shows how to convert Fischer projections to wedge dash representations.

Figure \(\PageIndex{4}\) shows an interactive iCn3D model of D-glucose in a linear form.
Orient the molecule as shown in Figure \(\PageIndex{5}\) below, with the carbonyl oxygen pointed to the far right, and compare it to the orientation shown in Figure \(\PageIndex{5}\) to reinforce your understanding of Fischer and wedge/dash projections.
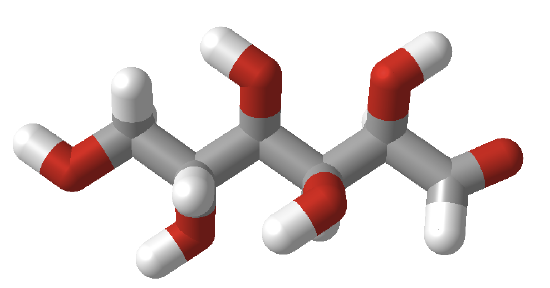
Figure \(\PageIndex{5}\):
Cyclic forms can be drawn either as the Haworth projections, which show the molecule as cyclic and planar with substituents above or below the ring, or the more plausible bent forms (showing glucose in the chair or boat conformations). β-D-glucopyranose is the only aldohexose that can be drawn with all its bulky substituents (OH and CH2OH) in equatorial positions, which probably accounts for its widespread prevalence in nature. Figure \(\PageIndex{6}\) shows four different representations of glucose.

Haworth projections are more realistic than the Fischer projections, but you should be able to draw both structures. Generally, if a substituent points to the right in the Fischer projection, it points down in the Haworth projection. If it points left, it points up. Generally, the OH on the α-anomer points down (αnts down) while on the β-anomer, it points up (βutterflies up) as illustrated in Figure \(\PageIndex{7}\)

In the Haworth projections, the bulky R group of the next carbon after the carbon whose OH group was the nucleophile for ring formation is pointed up if the OH engaged in the attack was on the right-hand side in the straight chain Fischer diagram (as in α-D-glucopyranose above, when the CH2OH group is up). Conversely, it is down if the OH engaged in the attack was on the left-hand side in the straight chain Fischer diagram (as in α-D-galactofuranose above, when the (CHOH)CH2OH group is down). The rest of the OH groups still follow the simple rule that if they point to the right in the Fischer projection, they point down in the Haworth projection.
The Fischer structures of the most common monosaccharides (other than glyceraldehyde and dihydroxyacetone), which you will encounter most frequently, are shown in Figure \(\PageIndex{8}\).

Figure \(\PageIndex{8}\): Most common monosaccharides discussed in this book
The mirror image of D-Glc is L-Glc. The D- and L- designations refer to the center of asymmetry most remote from the aldehyde or ketone. By convention, all chiral centers are related to D-glyceraldehyde, so sugar isomers related to D-glyceraldehyde at their last asymmetric center are D sugars.
Figure \(\PageIndex{9}\) shows multiple renderings of common hexoses.

Isomers
Sugars can be configurational (interconverted only by breaking covalent bonds) or conformational isomers. Figure \(\PageIndex{10}\) reviews different configurational isomers.

The configurational isomers include enantiomers (stereoisomers that are mirror images of each other), diastereomers (stereoisomers that are not mirror images), epimers (diastereomers that differ at one stereocenter), and anomers (a particular form of stereoisomer, diastereomer, and epimer). Figure \(\PageIndex{11}\) below shows enantiomers, diastereomers, epimers, and anomers of 6-carbon sugars.

Sugars can also exist as conformational isomers, which interchange without breaking covalent bonds. These include chair and boat conformations of the cyclic sugars as shown in Figure \(\PageIndex{12}\).

Monosaccharide Derivatives
Many derivatives of monosaccharides are found in nature. These include
- oxidized forms in which the aldehyde and/or alcohol functional groups are oxidized to carboxylic acids
- phosphorylated forms in which phosphates are transferred from ATP to form phosphoester derivatives
- amine derivatives such as glucosamine or galactosamine
- acetylated amine derivatives such as N-Acetyl-GlcNAc (GlcNAc) or GalNAc
- lactone forms (intramolecular esters) in which an OH group attacks a carbonyl C that was previously oxidized to a carboxylic acid
- condensation products of sugar derivatives with lactate (CH3CHOHCO2-) and pyruvate (CH3COCO2- ), both from the glycolytic pathway, to form muramic acid and neuraminic acids (also called sialic acids), respectively.
Figure \(\PageIndex{13}\) some simple monosaccharide derivatives.

Figure \(\PageIndex{14}\) shows some additional oxidative derivatives of glucose shown in Fischer projections.

Other important derivatives of monosaccharides are sialic acids. N-acetylmuramic acid, found in bacterial cell walls, consists of GlcNAc in an ether link at C3 with lactate, while N-acetylneuraminic acid results from an intramolecular cyclization of a condensation product of ManNAc and pyruvate. These sialic acids are shown in Figure \(\PageIndex{15}\).

Sugars are very complex, as their linkages and substituents are highly diverse. Figure \(\PageIndex{16}\) shows differences in sialic acids between humans and chimps.

What happens when non-vegan humans eat animal products (meat, milk) with N-glycoyl neuraminic acids (Neu5Gc)? Some get incorporated into human membrane glycans. Sialic acids on surface proteins can serve as "receptors" that bind self-cells and foreign cells or proteins that have evolved to bind them. A toxin, SubAB, secreted by E. Coli 0157, can bind Neu5Gc. Hence, eating meat products can make us more susceptible to bacteria that recognize Neu5Gc.
Formation of Hemiacetals, Acetals, and Disaccharides
Monosaccharides that contain an aldehyde can cyclize via an intramolecular nucleophilic attack by an OH group on the carbonyl carbon, forming a hemiacetal. In the past, the group was called a hemiketal if the attack was on a ketone, but now it is also called a hemiacetal. On the addition of acid (which protonates the anomeric OH, forming water as a potential leaving group), another alcohol can add, forming an acetal with water leaving. These reactions are shown in Figure \(\PageIndex{17}\).

If the other alcohol is a second monosaccharide, a disaccharide results. The acetal linkage between the two monosaccharides is called a glycosidic linkage. If the link between the two sugars involves an anomeric carbon, the newly formed OH group at the link can be designated either as α (if the O on the carbon involved in the glycosidic link is pointing down) or β (if the O is pointing up). For a 2-2 link between hexoses (i.e., between two non-anomeric carbons, the α/β designation is not used. Since sugars contain so many OH groups, which can act as the "second" alcohol in acetal formation, the links between sugars can be quite diverse. These include α and β forms of 1-2, 1-3, 1-4, 1-5, 1-6, etc. links. Here are examples of disaccharides:
- maltose: Glc(α 1,4)Glc, which can be considered a disaccharide hydrolysis product of the polysaccharide glycogen or starch (discussed in the section)
- cellobiose: Glc(Glc(α 1,4)Glc 1,4)Glc, which can be considered a disaccharide hydrolysis product of cellulose
- lactose: Gal(β 1,4)Glc, also known as milk sugar
- sucrose: Glc(α 1,2)Fru. Since fructose is attached through the anomeric OH of this ketose, it is not in equilibrium with its straight-chain keto form; hence, sucrose is a nonreducing sugar. Note also that since the anomeric C-OH of each sugar is used, the α/β designation in the disaccharide is used. Hence, sucrose would be abbreviated as Glc(α1,2β)Fru
Figure \(\PageIndex{18}\) illustrates the differences between lactose and sucrose. Note that the β-D-fructofuranose ring is flipped (left to right as in turning one of your hands over) compared to Figure \(\PageIndex{16}\).

Acetal links between sugars in glycans can be hydrolyzed by water (catalyzed by H+), as with the other key biological polymers, proteins, and nucleic acids.
The disaccharides described above, linked through a 1,4 linkage, are called reducing sugars because they can act as reducing agents in reactions in which they are oxidized. For example, in lactose, since galactose is attached to glucose through the OH on C4, the anomeric glucose carbon, C1, could revert to the noncyclic aldehyde form. This aldehyde is susceptible to oxidation by reagents (Benedict's Solution with citrate, Fehling's reagent with tartrate), as these reagents are subsequently reduced. In both reagents, reducing sugars reduce a basic blue solution of CuSO4 (Cu2+) to a brick-red precipitate of Cu2O (Cu1+). Sugars (monosaccharides, disaccharides and polysaccharides) that have a potentially open aldehyde at C1 or have an α-hydroxymethyl ketone group which can isomerize to an aldehyde under basic conditions (such as fructose) are called reducing sugars. These oxidizing agents are mild and react with aldehydes, but not with ketones.
A monosaccharide, disaccharide, or even polysaccharide with at least one hemiacetal link (for instance, lactose) is a reducing sugar, as the monomer with the cyclic hemiacetal can reversibly open to form an aldehyde. However, if the only links in sugars are full acetals (such as in sucrose, when the link is between the two anomeric carbons), the sugar is not reducing.
Alpha-gal syndrome (AGS) is a relatively new disease caused by a tick bite. Tick saliva contains the disaccharide galactose-α-1,3-galactose (alpha-gal). After a tick bite, people develop an immune response to the disaccharide through IgE antibodies. Further bites could cause a mild rash or a reaction as severe as an anaphylactic response.
Red meat contains the disaccharide, but it is not found in fish, birds, or people. Hence, people who mount a strong IgE response to the disaccharide can also elicit the same response when they eat red meat or even drink cow's milk. It is estimated that up to 450,000 people in the US may develop serious and even life-threatening symptoms after eating red meat.
The structure of Gal(α1,3)Gal is shown in Figure \(\PageIndex{19}\) below.

Figure \(\PageIndex{19}\): The structure of the disaccharide Gal(α1,3)Gal
Figure \(\PageIndex{20}\) shows an interactive iCn3D model of α-D-galactosyl-(1,3)-α-D-galactose (PubChem 9840966)

Figure \(\PageIndex{20}\): α-D-galactosyl-(1,3)-α-D-galactose (PubChem 9840966). (Copyright; author via source).
Click the image for a popup or use this external link: https://www.ncbi.nlm.nih.gov/Structu...5c4f77d7102ad6
Figure \(\PageIndex{21}\) shows an interactive iCn3D model of human anti-alpha-galactosyl antibody complex (7UEN)
.png?revision=1&size=bestfit&width=393&height=255)
Figure \(\PageIndex{20}\): Human anti-alpha-galactosyl antibody complex (7UEN). (Copyright; author via source).
Click the image for a popup or use this external link: https://www.ncbi.nlm.nih.gov/Structu...57bc3eaca2a3f0
This antibody in this complex is an IgG, not an IgE. The Fab light chain fragment of the antibody is in magenta, and the heavy chain Fab fragment in blue. Two side chains, H27 and S91, show interactions with the disaccharide.
Here is another iCn3D link that shows more interactions: https://www.ncbi.nlm.nih.gov/Structure/icn3d/share2.html?22f544b2836aadd2332a8ab3edd98be1. Gal residues are shown as yellow cubes in the SNFG color scheme. The abbreviation for Gal in alpha linkage in the PDB is GLA.
Summary
(Summary written by Claude, Sonnet 4.6, Anthropic)
This chapter introduces the structural chemistry of carbohydrates — the most chemically complex class of biological macromolecular building blocks — covering monosaccharide structure, stereochemistry, ring formation, chemical derivatives, and the formation and properties of glycosidic bonds in disaccharides.
Carbohydrates (sugars, glycans) are polyhydroxy aldehydes (aldoses) or polyhydroxy ketones (ketoses) that range from simple trioses to large polysaccharides. The name reflects the empirical formula of simple sugars like glucose (C₆H₁₂O₆ = C₆(H₂O)₆). Sugars are classified by the number of carbons (triose, tetrose, pentose, hexose) and by whether the carbonyl is an aldehyde (aldose) or ketone (ketose). The D/L designation derives from the configuration of the asymmetric center farthest from the carbonyl group relative to D-glyceraldehyde, the reference compound for all sugar stereochemistry. All common biological monosaccharides are D-sugars.
Structural representations of sugars demand facility with multiple conventions. In Fischer projections (used for open-chain forms), the horizontal bonds project toward the viewer and the vertical bonds project away; substituents to the right of the chain in a Fischer projection appear below the ring in Haworth projections. Fischer projections can be converted to wedge-dash drawings by orienting the carbon chain vertically with the carbonyl at top, then determining the relative positions of OH groups at each center. Haworth projections show the cyclic form as a flat ring, with the anomeric OH pointing down for α-anomers (αnts down) and up for β-anomers (βutterflies up); the bulky CH₂OH group at C5 points up in D-sugars. More realistic chair conformations show the actual three-dimensional geometry: β-D-glucopyranose is uniquely stable because all four OH groups and the CH₂OH group occupy equatorial positions in the ⁴C₁ chair, likely explaining glucose's predominance in biology.
Ring formation proceeds by intramolecular hemiacetal formation: a hydroxyl group on C5 (forming a six-membered pyranose ring) or C4 (forming a five-membered furanose ring) attacks the carbonyl carbon in a nucleophilic addition reaction. The new stereocenters at the anomeric carbon give α- and β-anomers that interconvert in solution via the open-chain aldehyde form—a process called mutarotation. Glucose in solution is ~64% β-pyranose and ~36% α-pyranose with only ~0.002% open-chain form; fructose is 67% pyranose and 33% furanose; ribose is 75% furanose. In polysaccharides, glucose is exclusively pyranose while fructose and ribose are furanose.
Stereoisomerism in sugars takes multiple forms requiring precise terminology. Enantiomers are mirror-image pairs (D-Glc and L-Glc) that differ at every stereocentre and are interconvertible only by breaking bonds. Diastereomers are non-mirror-image stereoisomers (e.g., glucose and galactose, which differ at C4), and the subset differing at exactly one stereocentre are epimers. Anomers — the α and β forms at the anomeric carbon — are a special class of epimers that interconvert in solution without breaking the ring through mutarotation; all other epimers require bond breaking to interconvert. Conformational isomers (chair, boat, twist-boat conformations of pyranoses) interconvert without any bond breaking.
Monosaccharide derivatives greatly expand the chemical diversity of the glycome. Oxidation of the C1 aldehyde yields aldonic acids (e.g., gluconate); oxidation of the C6 primary alcohol yields uronic acids (e.g., glucuronate, a key component of proteoglycans); dual oxidation produces aldaric acids (e.g., glucarate). Phosphorylation at C1 or C6 by ATP-dependent kinases generates metabolic intermediates (glucose-6-phosphate, fructose-1,6-bisphosphate). Amination at C2 produces amino sugars (glucosamine, galactosamine), and N-acetylation of these amines gives GlcNAc and GalNAc — ubiquitous components of glycoproteins, proteoglycans, and bacterial cell walls. Condensation of GlcNAc with lactate gives N-acetylmuramic acid (MurNAc), the distinctive monosaccharide of bacterial peptidoglycan. Condensation of ManNAc with pyruvate and intramolecular cyclization gives N-acetylneuraminic acid (Neu5Ac, a sialic acid), a negatively charged nine-carbon sugar abundant on mammalian cell surfaces; the related Neu5Gc (N-glycolylneuraminic acid), present in many animals but not in humans, illustrates how minor sugar modifications create immunologically distinct surfaces — humans who eat red meat can incorporate Neu5Gc into their glycans and become susceptible to toxins (like SubAB from E. coli O157) that recognize it.
Glycosidic bond formation converts hemiacetals (reversible, open to ring-chain equilibrium) into full acetals (stable, nonhydrolyzable without acid or enzyme catalysis). Under acid conditions, the anomeric OH is protonated and departs as water, generating an oxocarbenium ion that is attacked by a second hydroxyl group (from another sugar or aglycon) to form the O-glycosidic bond. The diversity of glycosidic bonds — arising from the choice of which anomeric carbon, which configuration (α or β), and which OH on the second sugar serves as the acceptor — generates enormous structural diversity not possible in nucleic acids or proteins. Key disaccharides include maltose (Glc(α1→4)Glc, from starch/glycogen hydrolysis), cellobiose (Glc(β1→4)Glc, from cellulose hydrolysis — the single linkage difference from maltose determines whether the polymer is digestible or structural), lactose (Gal(β1→4)Glc, milk sugar), and sucrose (Glc(α1→2β)Fru, table sugar, a nonreducing sugar because both anomeric carbons are engaged in the glycosidic bond and neither can open to a free aldehyde). Reducing sugars — those with a free hemiacetal that can open to an aldehyde — reduce Cu²⁺ in Benedict's and Fehling's reagents, a reaction used clinically and diagnostically. The disaccharide Gal(α1→3)Gal (alpha-gal), present in tick saliva and red meat but absent from humans, causes alpha-gal syndrome: tick bites trigger IgE antibody production against this epitope, leading to delayed anaphylactic reactions to red meat in sensitized individuals, affecting potentially 450,000 Americans.