
14.2: Frog Embryology
- Page ID
- 5137

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)The Egg
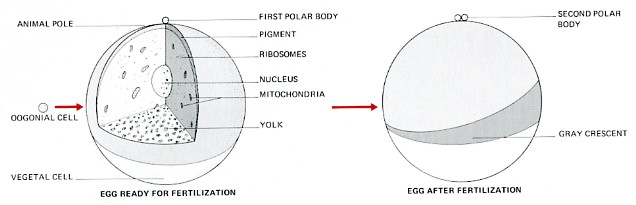
The frog egg is a huge cell; its volume is over 1.6 million times larger than a normal frog cell. During embryonic development, the egg will be converted into a tadpole containing millions of cells but containing the same amount of organic matter.
- The upper hemisphere of the egg — the animal pole — is dark.
- The lower hemisphere — the vegetal pole — is light.
- When deposited in the water and ready for fertilization, the haploid egg is at metaphase of meiosis II.
Fertilization

Entrance of the sperm initiates a sequence of events:
- Meiosis II is completed.
- The cytoplasm of the egg rotates about 30 degrees relative to the poles.
- In some amphibians (including Xenopus), this is revealed by the appearance of a light-colored band, the gray crescent.
- The gray crescent forms opposite the point where the sperm entered.
- It foretells the future pattern of the animal: its dorsal (D) and ventral (V) surfaces; its anterior (A) and posterior (P); its left and right sides.
- The haploid sperm and egg nuclei fuse to form the diploid zygote nucleus.
Cleavage
The zygote nucleus undergoes a series of mitoses, with the resulting daughter nuclei becoming partitioned off, by cytokinesis, in separate, and ever-smaller, cells. The first cleavage occurs shortly after the zygote nucleus forms. A furrow appears that runs longitudinally through the poles of the egg, passing through the point at which the sperm entered and bisecting the gray crescent. This divides the egg into two halves forming the 2-cell stage. The second cleavage forms the 4-cell stage. The cleavage furrow again runs through the poles but at right angles to the first furrow. The furrow in the third cleavage runs horizontally but in a plane closer to the animal than to the vegetal pole. It produces the 8-cell stage.

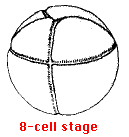
The next few cleavages also proceed in synchrony, producing a 16-cell and then a 32-cell embryo. However, as cleavage continues, the cells in the animal pole begin dividing more rapidly than those in the vegetal pole and thus become smaller and more numerous. By the next day, continued cleavage has produced a hollow ball of thousands of cells called the blastula. A fluid-filled cavity, the blastocoel, forms within it.
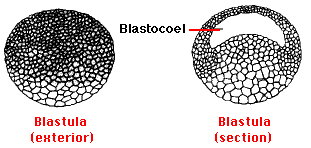
During this entire process there has been no growth of the embryo. In fact, because the cells of the blastula are so small, the blastula looks just like the original egg to the unaided eye. Not until the blastula contains some 4,000 cells is there any transcription of zygote genes. All of the activities up to now have been run by gene products (mRNA and proteins) deposited by the mother when she formed the egg.
Gastrulation
The start of gastrulation is marked by the pushing inward ("invagination") of cells in the region of the embryo once occupied by the middle of the gray crescent.
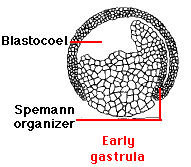

This produces an opening (the blastopore) that will be the future anus. a cluster of cells that develops into the Spemann organizer (named after one of the German embryologists who discovered its remarkable inductive properties).
As gastrulation continues, three distinct "germ layers" are formed:
- ectoderm
- mesoderm
- endoderm
Each of these will have special roles to play in building the complete animal. Some are listed in the table.
| Ectoderm | Mesoderm | Endoderm |
|---|---|---|
| skin | notochord | inner lining of gut, liver, pancreas |
| brain | muscles | inner lining of lungs |
| spinal cord | blood | inner lining of bladder |
| all other neurons | bone | thyroid and parathyroid glands |
| sense receptors | sex organs | thymus |

The Spemann organizer (mostly mesoderm) will develop into the notochord, which is the precursor of the backbone and induce the ectoderm lying above it to begin to form neural tissue instead of skin. This ectoderm grows up into two longitudinal folds, forming the neural folds stage. In time the lips of the folds fuse to form the neural tube. The neural tube eventually develops into the brain and spinal cord.
Differentiation
Although the various layers of cells in the frog gastrula have definite and different fates in store for them, these are not readily apparent in their structure. Only by probing for different patterns of gene expression (e.g., looking for tissue-specific proteins) can their differences be detected. In due course, however, the cells of the embryo take on the specialized structures and functions that they have in the tadpole, forming neurons, blood cells, muscle cells, epithelial cells, etc., etc.
Growth
At the time the tadpole hatches, it is a fully-formed organism. However, it has no more organic matter in it than the original frog egg had. Once able to feed, however, the tadpole can grow. It gains additional molecules with which it can increase the number of cells that make up its various tissues.