
15.11.1: Innate Behavior
- Page ID
- 5763

Behavior is action that alters the relationship between an organism and its environment. Behavior may occur as a result of
- an external stimulus (e.g., sight of a predator)
- internal stimulus (e.g., hunger)
- or, more often, a mixture of the two (e.g., mating behavior)
It is often useful to distinguish between
- innate behavior = behavior determined by the "hard-wiring" of the nervous system. It is usually inflexible, a given stimulus triggering a given response. A salamander raised away from water until long after its siblings begin swimming successfully will swim every bit as well as they the very first time it is placed in the water. Clearly this rather elaborate response is "built in" in the species and not something that must be acquired by practice.
- learned behavior = behavior that is more or less permanently altered as a result of the experience of the individual organism (e.g., learning to play baseball well).
- However, careful analysis often reveals that any particular behavior is a combination of innate and learned components.
Examples of innate behavior:
- taxes
- reflexes
- instincts
Taxes
Some organisms respond to a stimulus by automatically moving directly toward or away from or at some defined angle to it. These responses are called taxes. They are similar to tropisms in plants except that actual locomotion of the entire organism is involved. Link to a detailed discussion.
Reflexes
The Withdrawal Reflex
When you touch a hot object, you quickly pull you hand away using the withdrawal reflex.
These are the steps:
- The stimulus is detected by receptors in the skin.
- These initiate nerve impulses in sensory neurons leading from the receptors to the spinal cord.
- The impulses travel into the spinal cord where the sensory nerve terminals synapse with interneurons.
- Some of these synapse with motor neurons that travel out from the spinal cord entering mixed nerves that lead to the flexors that withdraw your hand.
- Others synapse with inhibitory interneurons that suppress any motor output to extensors whose contraction would interfere with the withdrawal reflex.
The Stretch Reflex
The stretch reflex is examined (with a diagram) on a separate page. Link to it.
Instincts
Instincts are complex behavior patterns which, like reflexes, are inborn, rather inflexible, and valuable at adapting the animal to its environment. They differ from reflexes in their complexity. The entire body participates in instinctive behavior, and an elaborate series of actions may be involved.

The scratching behavior of a dog and a European bullfinch, shown here, is part of their genetic heritage. The widespread behavior of scratching with a hind limb crossed over a forelimb in common to most birds, reptiles, and mammals. (Picture courtesy of Rudolf Freund and Scientific American, 1958.) So instincts are inherited just as the structure of tissues and organs is. Another example.
- The African peach-faced lovebird carries nesting materials to the nesting site by tucking them in its feathers.
- Its close relative, the Fischer's lovebird, uses its beak to transport nesting materials.
- The two species can hybridize. When they do so, the offspring succeed only in carrying nesting material in their beaks. Nevertheless, they invariably go through the motions of trying to tuck the materials in their feathers first.
Foraging Behavior
Foraging for food is a crucial behavior for animals. Like all behavior, it requires the interaction of many components. Nonetheless, it turns out that in some animals, at least, foraging behavior can be altered by a single gene.
Drosophila melanogaster
The discovery of the genetic control of foraging in Drosophila began with the observations of Marla Sokolowski when she was an undergraduate biology student at the University of Toronto.
She noticed that Drosophila larvae, feeding in her culture vessels, displayed one of two distinct feeding patterns:
- "rovers" who moved rapidly over the surface of the culture medium
- "sitters who fed at a much more leisurely pace
She went on to find that this "bimodal" pattern of behavior
- continued when the larvae became adults
- was present in populations of wild fruit flies, not just in her laboratory colonies
After further years of research, she has shown that the behavior is under the control of a single gene, named for ("foraging"). Two alleles are present, at almost equal frequencies, that is, for is polymorphic.
- forR, which is dominant
- fors, the recessive
- About 70% of natural populations are rovers being either homozygous for forR or heterozygous (forR/fors).
- Sitters are homozygous for fors
Both alleles encode a PKG, a protein kinase (an enzyme that attaches phosphate groups to target proteins) that is activated by the "second messenger" cyclic GMP (cGMP). The enzyme encoded by the forR allele is more active than that encoded by fors. She and her colleagues have succeeded in inserting forR DNA into sitters who promptly become rovers.
Why this polymorphism? Why should alleles for two such different behaviors be maintained at such high frequency in the population?
One possible answer: it permits the population to thrive under varying food conditions:
- sitters are favored when food is abundant
- rovers are favored when competition for food is strong, such as in crowded cultures
Honeybees
Honeybees have their version of the for gene, called Amfor ("Apis mellifera for"). It, too, encodes a cGMP-dependent protein kinase (PKG). When worker bees first hatch, they remain in the hive tending to various housekeeping chores, such as feeding the larvae. But when they are 2–3 weeks old, they leave the hive and begin foraging for nectar and pollen. This change in behavior coincides with the increased expression of Amfor. When newly-hatched workers are treated with cGMP, the amount of PKG in their brains goes up and they quickly begin foraging instead of doing housekeeping.
Interaction of Internal and External Stimuli
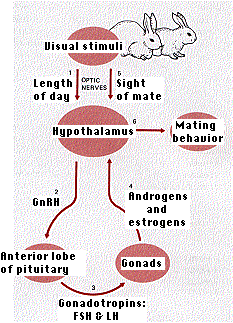
Instinctive behavior often depends on conditions in the internal environment. In many vertebrates courtship and mating behavior will not occur unless sex hormones (estrogens in females, androgens in males) are present in the blood. The target organ is a small region of the hypothalamus. When stimulated by sex hormones in its blood supply, the hypothalamus initiates the activities leading to mating. The level of sex hormones is, in turn, regulated by the activity of the anterior lobe of the pituitary gland.
The above figure outlines the interactions of external and internal stimuli that lead an animal, such as a rabbit, to see a sexual partner and mate with it.
Releasers of Instinctive Behavior
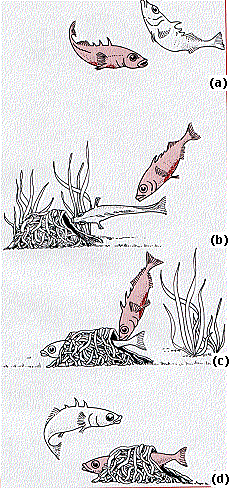
Once the body is prepared for certain types of instinctive behavior, an external stimulus may be needed to initiate the response. N. Tinbergen (who shared the 1973 Nobel Prize with Konrad Lorenz and Karl von Frisch) showed that the stimulus need not necessarily be appropriate to be effective.
- During the breeding season, the female three-spined stickleback normally follows the red-bellied male (a in the figure) to the nest that he has prepared.
- He guides her into the nest (b) and then
- prods the base of her tail (c).
- She then lays eggs in the nest.
- After doing so, the male drives her from the nest, enters it himself, and fertilizes the eggs (d).
- Although this is the normal pattern, the female will follow almost any small red object to the nest, and once within the nest, neither the male nor any other red object need be present.
- Any object touching her near the base of her tail will cause her to release her eggs.
It is as though she were primed internally for each item of behavior and needed only one specific signal to release the behavior pattern. For this reason, signals that trigger instinctive acts are called releasers. Once a particular response is released, it usually runs to completion even though the stimulus has been removed. One or two prods at the base of her tail will release the entire sequence of muscular actions involved in liberating her eggs.
Chemical signals (e.g., pheromones) serve as important releasers for the social insects: ants, bees, and termites. Many of these animals emit several different pheromones which elicit, for example, alarm behavior, mating behavior, and foraging behavior in other members of their species.
The mammary glands of domestic rabbit mothers emit a pheromone that releases immediate nursing behavior by their babies (pups). A good thing, too, as mothers devote only 5–7 minutes a day to feeding their pups so they had better be quick about it.
The studies of Tinbergen and others have shown that animals can often be induced to respond to inappropriate releasers. For example, a male robin defending its territory will repeatedly attack a simple clump of red feathers instead of a stuffed robin that lacks the red breast of the males.
Although such behavior seems inappropriate to our eyes, it reveals a crucial feature of all animal behavior: animals respond selectively to certain aspects of the total sensory input they receive. Animals spend their lives bombarded by myriad sights, sounds, odors, etc. But their nervous system filters this mass of sensory data, and they respond only to those aspects that the evolutionary history of the species has proved to be significant for survival.