
5.2: Central Metabolism
- Page ID
- 42513

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Learning Objectives
- Describe why glycolysis is not oxygen dependent
- Define and describe the net yield of three-carbon molecules, ATP, and NADH from glycolysis
- Explain how three-carbon pyruvate molecules are converted into two-carbon acetyl groups that can be funneled into the TCA cycle.
- Define and describe the net yield of CO2, GTP/ATP, FADH2, and NADH from the TCA cycle
- Explain how intermediate carbon molecules (precursor metabolites) of the TCA cycle can be used in anabolism
Extensive enzyme pathways exist for breaking down organic compounds to capture energy in ATP bonds. In almost all organisms, however, these catabolic pathways convert the initial substrates, such as carbohydrates or proteins, into metabolites that can enter glycolysis, the tricarboxylic acid cycle (TCA cycle; a.k.a. Krebs or citric acid cycle), or the pentose phosphate pathway (not covered here). These three pathways are referred to as "central metabolism" because almost all metabolic pathways, catabolic or anabolic, intersect with one or more of these pathways at some point. In addition to its role in catabolism, central metabolism produces intermediate molecules that are also used as building blocks for anabolism, the most critical of which are called precursor metabolites. Understanding these processes is important for several reasons. First, because the main metabolic processes involved are common to a wide range of chemoheterotrophic organisms, we can learn a great deal about human metabolism by studying metabolism in more easily manipulated bacteria like E. coli. Second, because animal and human pathogens are also chemoheterotrophs, learning about the details of metabolism in these bacteria, including possible differences between bacterial and human pathways, is useful for the diagnosis of pathogens as well as for the discovery of antimicrobial therapies targeting specific pathogens. Last, learning specifically about the pathways involved in chemoheterotrophic metabolism also serves as a basis for comparing other less familiar metabolic strategies used by microbes. Although the chemical source of electrons initiating electron transfer is different between chemoheterorophs and chemoautotrophs, the pathways of central metabolism are used by all types of organisms.
Glycolysis
For bacteria, eukaryotes, and most archaea, glycolysis is the most common pathway for the catabolism of glucose; it produces energy, reduced electron carriers, and precursor molecules for cellular metabolism. Every living organism carries out some form of glycolysis, suggesting this mechanism is an ancient universal metabolic process. The process itself does not use oxygen; however, glycolysis can be coupled with additional metabolic processes that are either aerobic or anaerobic. Glycolysis takes place in the cytoplasm of prokaryotic and eukaryotic cells. It begins with a single six-carbon glucose molecule and ends with two molecules of a three-carbon sugar called pyruvate. Pyruvate may be broken down further after glycolysis to harness more energy through aerobic or anaerobic respiration, but many organisms, including many microbes, may be unable to respire; for these organisms, glycolysis may be their only source of generating ATP.
The type of glycolysis found in animals and that is most common in microbes is the Embden-Meyerhof-Parnas (EMP) pathway, named after Gustav Embden (1874–1933), Otto Meyerhof (1884–1951), and Jakub Parnas (1884–1949). Glycolysis using the EMP pathway consists of three distinct phases (Figure \(\PageIndex{1}\)). The first part of the pathway, called the energy investment phase, uses energy from two ATP molecules to modify a glucose molecule so that the six-carbon sugar molecule can be split evenly into two phosphorylated three-carbon molecules called glyceraldehyde 3-phosphate (G3P). The splitting of the molecule itself is the second phase. The last part of the pathway, called the energy payoff phase, extracts energy by oxidizing G3P to pyruvate, producing four ATP molecules and reducing two molecules of NAD+ to two molecules of NADH, using electrons that originated from glucose.
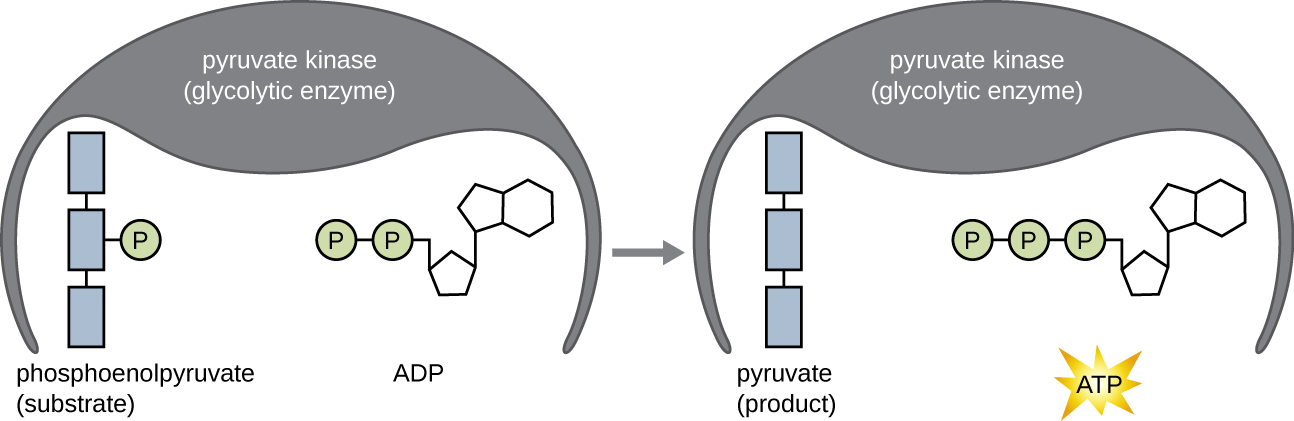
The ATP molecules produced during the energy payoff phase of glycolysis are formed by substrate-level phosphorylation (Figure \(\PageIndex{1}\)), one of two mechanisms for producing ATP. In substrate-level phosphorylation, a phosphate group is removed from an organic molecule and is directly transferred to an available ADP molecule, producing ATP. During glycolysis, high-energy phosphate groups from the intermediate molecules are added to ADP to make ATP.
Overall, in this process of glycolysis, the net gain from the breakdown of a single glucose molecule is:
- two ATP molecules
- two NADH molecule, and
- two pyruvate molecules.
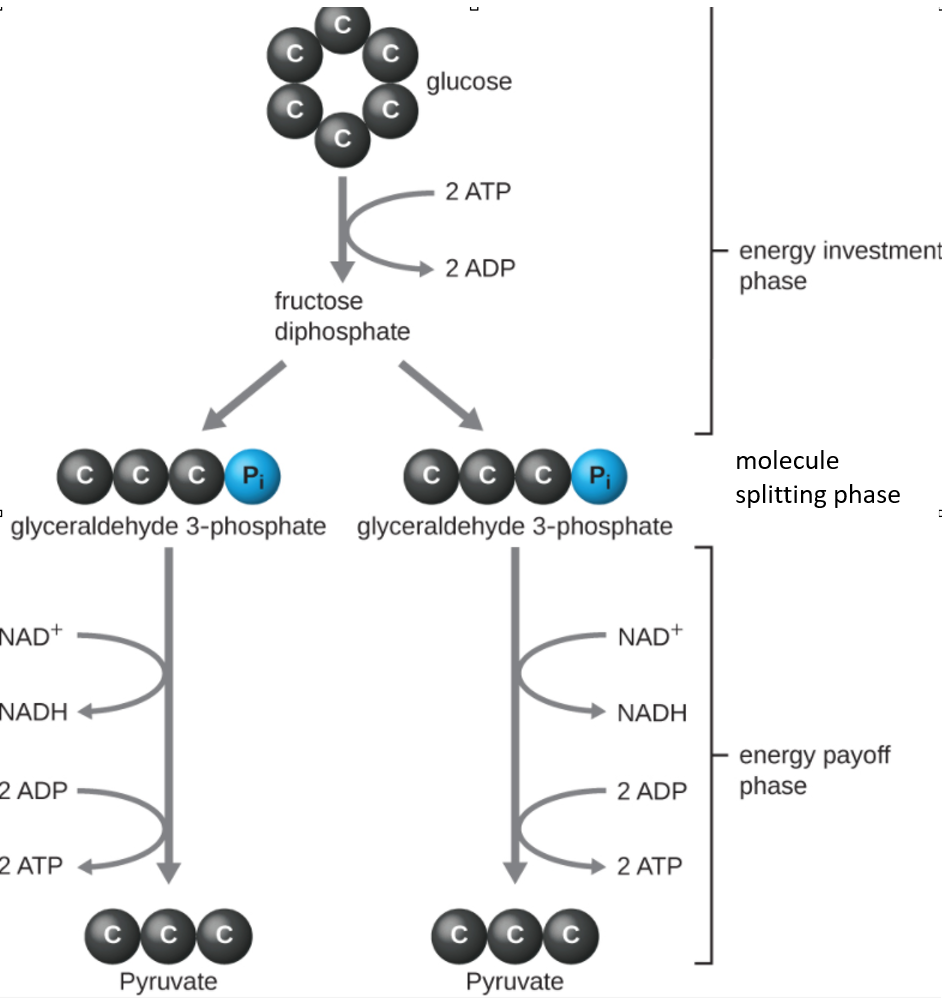
Figure \(\PageIndex{1}\): The energy investment phase of the Embden-Meyerhof-Parnas glycolysis pathway uses two ATP molecules to phosphorylate glucose, forming two glyceraldehyde 3-phosphate (G3P) molecules. The energy payoff phase harnesses the energy in the G3P molecules, producing four ATP molecules, two NADH molecules, and two pyruvates.
Other Glycolytic Pathways
When we refer to glycolysis, unless otherwise indicated, we are referring to the EMP pathway used by animals and many bacteria. However, some prokaryotes use alternative glycolytic pathways. One important alternative is the Entner-Doudoroff (ED) pathway, named after its discoverers Nathan Entner and Michael Doudoroff (1911–1975). Although some bacteria, including the opportunistic gram-negative pathogen Pseudomonas aeruginosa, contain only the ED pathway for glycolysis, other bacteria, like E. coli, have the ability to use either the ED pathway or the EMP pathway.
A third type of glycolytic pathway that occurs in all cells, which is quite different from the previous two pathways, is the pentose phosphate pathway (PPP) also called the phosphogluconate pathway or the hexose monophosphate shunt. Evidence suggests that the PPP may be the most ancient universal glycolytic pathway. The intermediates from the PPP are used for the biosynthesis of nucleotides and amino acids. Therefore, this glycolytic pathway may be favored when the cell has need for nucleic acid and/or protein synthesis, respectively. A discussion and illustration of the complete ED pathway and PPP with chemical structures and enzyme names appear in Appendix C.
Exercise \(\PageIndex{1}\)
When might an organism use the ED pathway or the PPP for glycolysis?
Transition Reaction, Coenzyme A, and the TCA Cycle
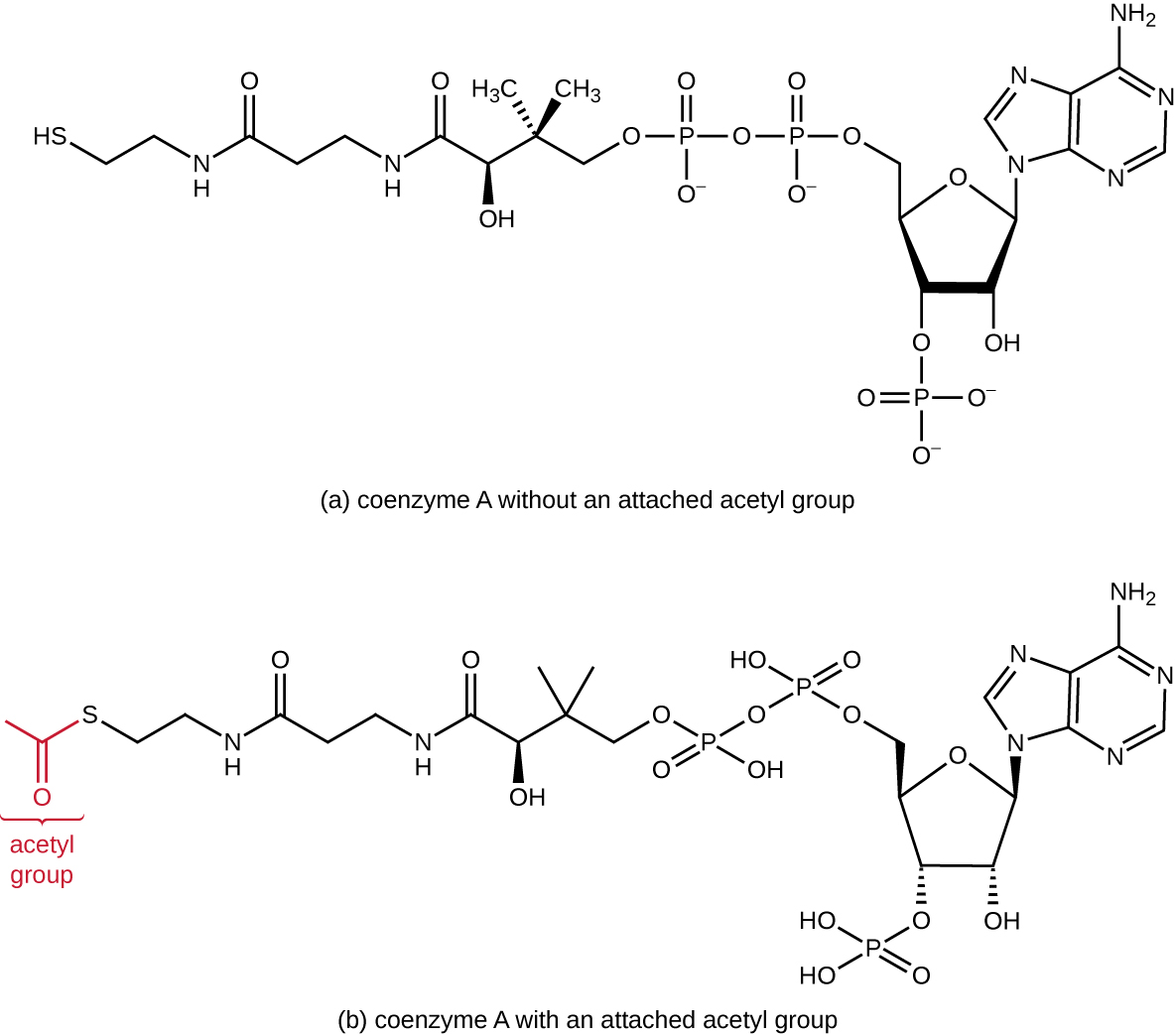
Glycolysis produces pyruvate, which can be further oxidized to capture more energy. For pyruvate to enter the next oxidative pathway, it must first be decarboxylated by the enzyme complex pyruvate dehydrogenase to a two-carbon acetyl group in the transition reaction, also called the bridge reaction (see Appendix C and Figure \(\PageIndex{3}\)). In the transition reaction, electrons are also transferred to NAD+ to form NADH. To proceed to the next phase of this metabolic process, the comparatively tiny two-carbon acetyl must be attached to a very large carrier compound called coenzyme A (CoA). The transition reaction occurs in the mitochondrial matrix of eukaryotes; in prokaryotes, it occurs in the cytoplasm because prokaryotes lack membrane-enclosed organelles.
The TCA cycle transfers remaining electrons from the acetyl group produced during the transition reaction to electron carrier molecules, thus reducing them. The TCA cycle also occurs in the cytoplasm of prokaryotes along with glycolysis and the transition reaction, but it takes place in the mitochondrial matrix of eukaryotic cells where the transition reaction also occurs. The TCA cycle is sometimes called the Krebs cycle after its discoverer, British scientist Hans Adolf Krebs (1900–1981) and is also called the citric acid cycle (a tricarboxylic acid) is the product of the first reaction of the cycle. Unlike glycolysis, the TCA cycle is a closed loop: The last part of the pathway regenerates the compound used in the first step (Figure \(\PageIndex{4}\)). The eight steps of the cycle are a series of chemical reactions that capture the two-carbon acetyl group (the CoA carrier does not enter the TCA cycle) from the transition reaction, which is added to four-carbon oxaloacetate in the TCA cycle, producing the six-carbon intermediate citric acid (giving the alternate name for this cycle). As one turn of the cycle returns to the starting point of the four-carbon intermediate, the cycle produces two CO2 molecules. Each time a CO2 molecule is released, the electrons from the bond holding the CO2 to the molecule (in the form of a carboxyl group) are captured on NAD+ to form NADH. In the regeneration phase of the TCA cylce, one ATP molecule (or an equivalent, such as guanosine triphosphate [GTP]) produced by substrate-level phosphorylation, one of FADH2, and one more NADH (for a total of three throughout the cylce) are produced.
Although many organisms use the TCA cycle as described as part of glucose metabolism, several of the intermediate compounds in the TCA cycle (referred to as precursor metabolites) can be used in synthesizing a wide variety of important cellular molecules, including amino acids, chlorophylls, fatty acids, and nucleotides; therefore, the cycle is both anabolic and catabolic (Figure \(\PageIndex{5}\)).
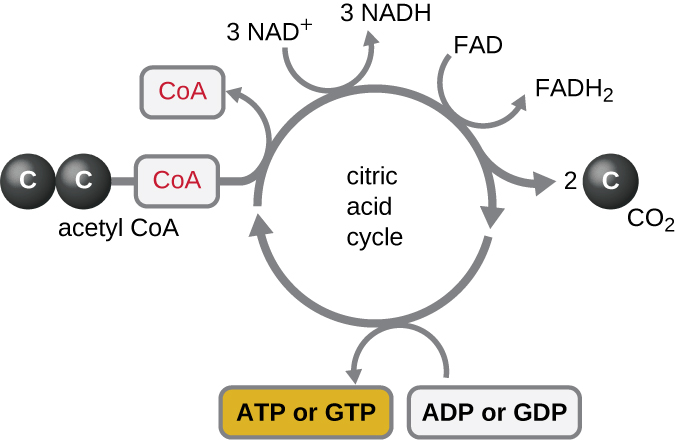
Figure \(\PageIndex{5}\): Many organisms use intermediates from the TCA cycle, such as amino acids, fatty acids, and nucleotides, as building blocks for biosynthesis.
Key Concepts and Summary
- Glycolysis is the first step in the breakdown of glucose, resulting in the formation of ATP, which is produced by substrate-level phosphorylation; NADH; and two pyruvate molecules. Glycolysis does not use oxygen and is not oxygen dependent.
- After glycolysis, a three-carbon pyruvate is decarboxylated to form a two-carbon acetyl group, coupled with the formation of NADH. The acetyl group is attached to a large carrier compound called coenzyme A.
- After the transition step, coenzyme A transports the two-carbon acetyl to the TCA cycle, where the two carbons enter the cycle. Per turn of the cycle, one acetyl group derived from glycolysis is further oxidized, producing three NADH molecules, one FADH2, and one ATP by substrate-level phosphorylation, and releasing two CO2molecules.
- The TCA cycle is used for other purposes. Many of the intermediates (precursor metabolites) are used to synthesize important cellular molecules, including amino acids, chlorophylls, fatty acids, and nucleotides.
Contributors and Attributions
Nina Parker, (Shenandoah University), Mark Schneegurt (Wichita State University), Anh-Hue Thi Tu (Georgia Southwestern State University), Philip Lister (Central New Mexico Community College), and Brian M. Forster (Saint Joseph’s University) with many contributing authors. Original content via Openstax (CC BY 4.0; Access for free at https://openstax.org/books/microbiology/pages/1-introduction)