
22.1: What is Biodiversity?
- Page ID
- 84101
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Genetic diversity, ecosystem diversity, and human-derived diversity are measures of biodiversity that currently define life on earth.
Key Points
- A genus with a high variety of species will have more genetic diversity; the most genetically-diverse species will have the greatest potential for evolution and preservation.
- The loss of ecosystem diversity results in the loss of interactions between species, unique features of co-adaptation, and biological productivity.
- Human-generated species diversity has decreased due to migration, market forces, and agriculture.
- Humans have only been able to estimate the number of species that inhabit Earth; this estimate only accounts for 20 percent of predicted species on the planet.
Key Terms
- genetic diversity: variety of genes in a species or other taxonomic group or ecosystem; can refer to allelic diversity or genome-wide diversity
- ecosystem diversity: variety of ecosystems in a biosphere or the variety of species and ecological processes that occur in different physical settings
- chemical diversity: variety of metabolic compounds in an ecosystem
Types of Biodiversity
Scientists generally accept that the term biodiversity describes the number and kinds of species in a location or on the planet. Species can be difficult to define, but most biologists still feel comfortable with the concept and are able to identify and count eukaryotic species in most contexts. Biologists have also identified alternate measures of biodiversity, some of which are important for planning how to preserve biodiversity.
Genetic diversity is one of those alternate concepts. Genetic diversity or variation is the raw material for adaptation in a species. A species’ future potential for adaptation depends on the genetic diversity held in the genomes of the individuals in populations that make up the species. The same is true for higher taxonomic categories. A genus with very different types of species will have more genetic diversity than a genus with species that look alike and have similar ecologies. If there were a choice between one of these genera of species being preserved, the one with the greatest potential for subsequent evolution is the most genetically-diverse one. It would be ideal not to have to make such choices, but, increasingly, this may be the norm.
Many genes code for proteins, which in turn carry out the metabolic processes that keep organisms alive and reproducing. Genetic diversity can be measured as chemical diversity in that different species produce a variety of chemicals in their cells, both the proteins as well as the products and by-products of metabolism. This chemical diversity has potential benefits for humans as a source of pharmaceuticals, so it provides one way to measure diversity that is important to human health and welfare.
Humans have generated diversity in domestic animals, plants, and fungi. This diversity is also suffering losses because of migration, market forces, and increasing globalism in agriculture, especially in heavily-populated regions such as China, India, and Japan. The human population directly depends on this diversity as a stable food source; its decline is troubling to biologists and agricultural scientists.
It is also useful to define ecosystem diversity: the number of different ecosystems on the planet or in a given geographic area. Whole ecosystems can disappear even if some of the species might survive by adapting to other ecosystems. The loss of an ecosystem means the loss of interactions between species, the loss of unique features of co-adaptation, and the loss of biological productivity that an ecosystem is able to create. An example of a largely-extinct ecosystem in North America is the prairie ecosystem. Prairies once spanned central North America from the boreal forest in northern Canada down into Mexico. Now, they have mostly disappeared, replaced by crop fields, pasture lands, and suburban sprawl. Many of the species survive, but the hugely-productive ecosystem that was responsible for creating the most productive agricultural soils is now gone. As a consequence, soils are disappearing or must be maintained at greater expense.

Figure \(\PageIndex{1}\): Ecosystem diversity - The variety of ecosystems on earth, from (a) coral reef to (b) prairie, enables a great diversity of species to exist.
Case Study Modified from Armstrong, C. G., Miller, J. E., McAlvay, A. C., Ritchie, P. M., & Lepofsky, D. (2021). Historical indigenous land-use explains plant functional trait diversity. Ecology and Society. 26: 6. This article was published under CC BY 4.0.
In addition to species diversity, ecologists are often interested in the trait or functional diversity of a community. A trait is simply any morphological, physiological or phenological feature measurable at the individual level (Reiss et al., 2009). Functional traits are those that define species in terms of their ecological roles - how they interact with the environment and with other species (Diaz & Cabido, 2001).
Functional diversity is a biodiversity measure based on functional traits of the species present in a community. In ocean phytoplankton, for example, these traits usually include body size, tolerance and sensitivity to environmental conditions, motility, shape, and N-fixation ability (Reynolds et al., 2002; Weithoff G., 2003). In terrestrial plant communities, researchers have included more complex traits like rates of growth, nutrient requirements and water uptake (Walker & Langridge, 2002; Barnett et al, 2007). Functional traits are a critical tool for understanding ecological communities because they give insights into community assembly processes as well as potential species interactions and other ecosystem functions. Because there are a greater variety of “roles” being played in a system with higher functional diversity, this measure of diversity has often been linked to higher ecosystem productivity and stability.
Human land-use legacies have long-term effects on plant community composition and ecosystem function and on the diversity of functional traits. Armstrong et al. (2021) studied how plant functional trait distributions and functional diversity are affected by ancient and historical Indigenous forest management in the Pacific Northwest.

Figure \(\PageIndex{2}\): The Village complex of Dałk Gyilakyaw consists of three discrete villages and is the ancestral home of Gitsm’geelm (Ts’msyen) people. Note the dramatic vegetation change between the forest garden and encroaching conifers (“periphery forests”). Photograph: S. Carroll.

Figure \(\PageIndex{3}\): Total Species Richness and Species Richness by Lifeform. Richness is indicated overall between forest gardens and periphery forests (averaged across the four of the study areas) and among the three growth forms (trees, shrubs and herbs).
For their research into plant functional diversity, Armstrong et al. (2021) compared forest garden ecosystems - managed perennial fruit and nut communities associated exclusively with archaeological village sites - with surrounding periphery conifer forests. To characterize the functional diversity of understory plant communities, they focused on four functional traits: seed mass, shade tolerance, pollination syndrome, and dispersal syndrome. These traits represent important axes of plant life-history variation and can also have important consequences for ecosystem functioning, while also having relevance to ethnobotanical plant uses (Pérez-Harguindeguy et al., 2013). For example, plants with animal-dispersed seeds may be able to disperse long distances and may also contribute to wildlife habitat by providing edible fruits; these plants are also more likely to be eaten by people.
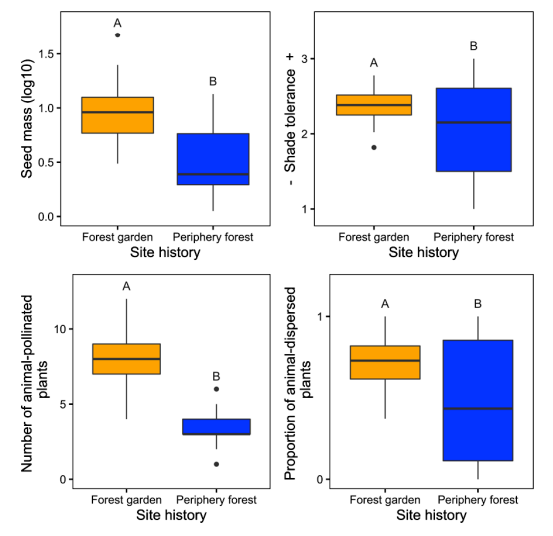
Figure \(\PageIndex{4}\): Functional Trait Measures between Forest Gardens and Periphery Forests. Comparisons of average seed mass, shade tolerance, pollination syndrome, and dispersal syndrome traits for herbs and shrubs across forest gardens and peripheral forests — all are significantly higher in the forest gardens.
Armstrong et al. (2021) found that forest gardens have substantially greater plant and functional trait diversity than periphery forests, even more than 150 years after management ceased. Forests managed by Indigenous peoples in the past now provide diverse resources and habitat for animals and other pollinators and are more rich than naturally forested ecosystems. Although ecological studies rarely incorporate Indigenous land-use legacies, the positive effects of Indigenous land use on contemporary functional and taxonomic diversity found by Armstrong et al. (2021) suggest that Indigenous management practices are tied to ecosystem health and resilience.
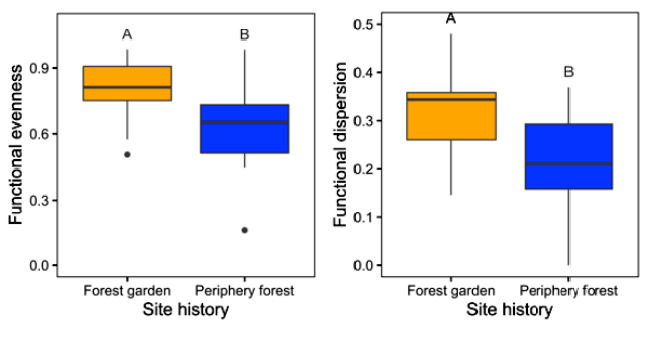
Figure \(\PageIndex{5}\): Functional Diversity Measures between Forest Gardens and Periphery Forests. Functional evenness (the evenness of functional trait distribution in niche space; Villéger et al. 2008) and functional dispersion (the average distance to the abundance-weighted centroid of functional trait values; Laliberté, and Legendre 2010) were significantly greater in forest gardens as compared to periphery forests. Comparisons of functional diversity at forest gardens and peripheral forests.
References
Barnett, A.J., & Beisner, B.E. (2007). Zooplankton biodiversity and lake tropic state: Explanations invoking resource abundance and distribution. Ecology, 88, pp. 1675-1686
Diaz, S., & Cabido, M. (2001). Vive la difference: Plant functional diversity matters to ecosystem processes. Trends in Ecology and Evolution, 16, pp. 646-655
Pérez-Harguindeguy N., Díaz, S., Garnier, E., Lavorel, S., Poorter, H., Jaureguiberry, P., Bret-Harte, M.S., Cornwell, W.K., Craine, J.M., Gurvich, D.E., Urcelay, C., Veneklaas, E.J., Reich, P.B., Poorter, L., Wright, I.J., Ray, P., Enrico, L., Pausas, J.G., de Vos, N. Buchmann, A.C., Funes, G., Quétier, F., Hodgson, J.G., Thompson, K., Morgan, H.D., ter Steege, H., van der Heijden, M.G.A., Sack, L., Blonder, B., Poschlod, P., Vaieretti, M.V., Conti, G., Staver, A.C., Aquino, S., & Cornelissen, J.H.C. (2013). New handbook for standardised measurement of plant functional traits worldwide. Australian Journal of Botany, 61, pp. 167-234.
Reiss, J., Bridle, J.R., Montoya, J.M. and Woodward, G. (2009). Emerging horizons in biodiversity and ecosystem functioning research. Trends Ecol. Evol., 24, pp. 505-514
Reynolds, C.S., Huszar, V., Kruk, C., Naselli-Flores, L. and Melo, S. (2002). Towards a functional classification of the freshwater phytoplankton. Journal of Plankton Research, 24, pp. 417–428
Villéger, S., Mason, N.W., & Mouillot, D. (2008). New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology, 89, pp. 2290-2301.
Walker, B.H., & Langridge, J.L. (2002). Measuring functional diversity in plant communities with mixed lifeforms: A problem of hard and soft attributes. Ecosystems, 5, pp. 529–538
Weithoff, G. (2003). The concept of ‘plant functional types’ and ‘functional diversity’ in lake phytoplankton – new understanding of phytoplankton ecology? Freshwater Biology, 48, pp. 1669–1675
Contributors and Attributions
Written and modified by A. Wilson and N. Gownaris (Gettysburg College) from the following open-access sources