
30.18: Plant Sensory Systems and Responses - The Phytochrome System and Red Light Response
- Page ID
- 13766

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)- Explain the response of the phytochrome system to red/far-red light
The phytochromes are a family of chromoproteins with a linear tetrapyrrole chromophore, similar to the ringed tetrapyrrole light-absorbing head group of chlorophyll. Phytochromes have two photo-interconvertible forms: Pr and Pfr. Pr absorbs red light (~667 nm) and is immediately converted to Pfr. Pfr absorbs far-red light (~730 nm) and is quickly converted back to Pr. Absorption of red or far-red light causes a massive change to the shape of the chromophore, altering the conformation and activity of the phytochrome protein to which it is bound. Pfr is the physiologically-active form of the protein; exposure to red light yields physiological activity in the plant. Exposure to far-red light converts the Pfr to the inactive Pr form, inhibiting phytochrome activity. Together, the two forms represent the phytochrome system.
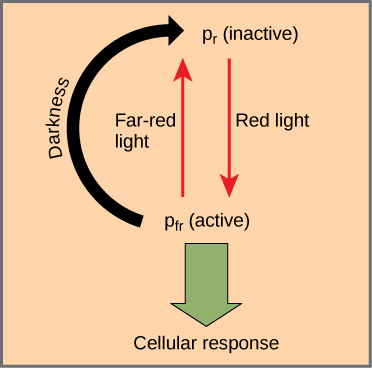
The phytochrome system acts as a biological light switch. It monitors the level, intensity, duration, and color of environmental light. The effect of red light is reversible by immediately shining far-red light on the sample, which converts the chromoprotein to the inactive Pr form. Additionally, Pfr can slowly revert to Pr in the dark or break down over time. In all instances, the physiological response induced by red light is reversed. The active form of phytochrome (Pfr) can directly activate other molecules in the cytoplasm, or it can be trafficked to the nucleus, where it directly activates or represses specific gene expression.
The Phytochrome System and Growth
Plants use the phytochrome system to grow away from shade and toward light. Unfiltered, full sunlight contains much more red light than far-red light. Any plant in the shade of another plant will be exposed to red-depleted, far-red-enriched light because the other plant has absorbed most of the other red light. The exposure to red light converts phytochrome in the shaded leaves to the Pr (inactive) form, which slows growth. The leaves in full sunlight are exposed to red light and have activated Pfr, which induces growth toward sunlit areas. Because competition for light is so fierce in a dense plant community, those plants who could grow toward light the fastest and most efficiently became the most successful.
The Phytochrome System in Seeds
In seeds, the phytochrome system is used to determine the presence or absence of light, rather than the quality. This is especially important in species with very small seeds and, therefore, food reserves. For example, if lettuce seedlings germinated a centimeter under the soil surface, the seedling would exhaust its food resources and die before reaching the surface. A seed will only germinate if exposed to light at the surface of the soil, causing Pr to be converted to Pfr, signaling the start of germination. In the dark, phytochrome is in the inactive Pr form so the seed will not germinate.
Photoperiodism
Plants also use the phytochrome system to adjust growth according to the seasons. Photoperiodism is a biological response to the timing and duration of dark and light periods. Since unfiltered sunlight is rich in red light, but deficient in far-red light, at dawn, all the phytochrome molecules in a leaf convert to the active Pfr form and remain in that form until sunset. Since Pfr reverts to Pr during darkness, there will be no Pfr remaining at sunrise if the night is long (winter) and some Pfr remaining if the night is short (summer). The amount of Pfr present stimulates flowering, setting of winter buds, and vegetative growth according to the seasons.
In addition, the phytochrome system enables plants to compare the length of dark periods over several days. Shortening nights indicate springtime to the plant; lengthening nights indicate autumn. This information, along with sensing temperature and water availability, allows plants to determine the time of the year and adjust their physiology accordingly. Short-day (long-night) plants use this information to flower in the late summer and early fall when nights exceed a critical length (often eight or fewer hours). Long-day (short-night) plants flower during the spring when darkness is less than a critical length (often 8 to 15 hours). However, day-neutral plants do not regulate flowering by day length. Not all plants use the phyotochrome system to adjust their physiological responses to the seasons.
Key Points
- Exposure to red light converts the chromoprotein to the functional, active form (Pfr), while darkness or exposure to far-red light converts the chromophore to the inactive form (Pr).
- Plants grow toward sunlight because the red light from the sun converts the chromoprotein into the active form (Pfr), which triggers plant growth; plants in shade slow growth because the inactive form (Pr) is produced.
- If seeds sense light using the phytochrome system, they will germinate.
- Plants regulate photoperiodism by measuring the Pfr/Pr ratio at dawn, which then stimulates physiological processes such as flowering, setting winter buds, and vegetative growth.
Key Terms
- phytochrome: any of a class of pigments that control most photomorphogenic responses in higher plants
- chromophore: the group of atoms in a molecule in which the electronic transition responsible for a given spectral band is located
- photoperiodism: the growth, development and other responses of plants and animals according to the length of day and/or night