
19.1.10: Invertebrates
- Page ID
- 5958

The Origin and Evolution of Animals (Metazoa)
We do not yet know from what group(s?) of eukaryotes the animals evolved. It occurred in Precambrian times. Before the Cambrian was far along, most of the animal phyla had appeared. So each of the phyla described in this section has had a long, independent history. The rapid (geologically speaking!) diversification of the animals has made it difficult to establish the genealogical relationships between them — even using molecular data. Our best guesses are shown in the cladogram below..
Sponges (Phylum Porifera)
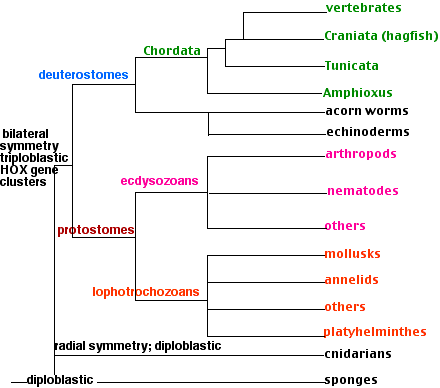
Sponges are sessile, spending their lives anchored to a solid surface underwater. Most are marine although some live in fresh water. Diploblastic; that is, the body wall is made of two layers of cells with a jellylike mesoglea between them. The body wall is perforated with pores (hence the name Porifera) through which water containing food particles is filtered. The water is drawn in through the pores by collar cells like those found in choanoflagellates. Some sponges can process a volume of water more than 100,000 times their own volume in the course of a day!
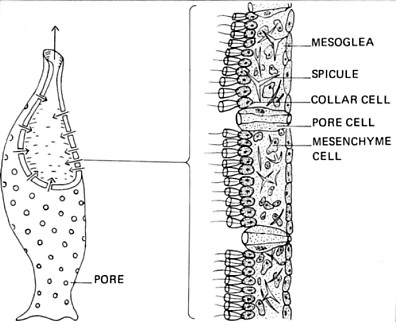
Sponges are dispersed by small, free-swimming larvae. There are about 10,000 species known. Sponges are probably the most ancient of today's invertebrates, their fossils appear in the geological record as far back as 635 million years. Despite their simple body plan, sequencing shows that their genome (> 18,000 genes) contains many genes homologous to those found in much more complex animals.
Cnidarians (Phylum Cnidaria)
Characteristics:
- diploblastic; two layers of cells - ectoderm and endoderm - with a jellylike mesoglea between them;
- predominantly radial symmetry: body parts (e.g., tentacles) arranged in whorls. However, in some sea anemones, there is only one plane through the tubular body that divides it into two mirror-image halves; thus revealing bilateral symmetry.
- cnidoblasts: specialized cells that secrete a stinging capsule called a nematocyst.
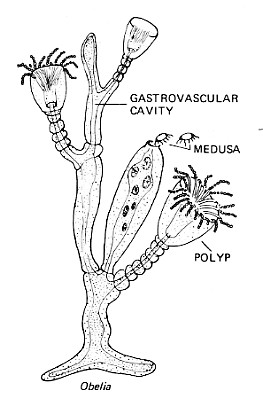
- Food is taken through a mouth into the gastrovascular cavity. The cavity is also called a coelenteron and for many years the name of this phylum was Coelenterata. There is no anus.
- Sexual reproduction produces a free-swimming, ciliated larva called a planula.
- The phylum contains about 10,000 species distributed in 3 classes:
- Hydrozoa Although the freshwater hydra is a much-studied representative, it is not typical of the class.
Most members are- marine
- colonial
- produce two body forms: the sessile polyp (like the hydra) and the free-floating medusa (which disperses the species)
- Scyphozoa Jellyfishes (the medusa stage is dominant). The jelly of the medusa is a much-enlarged mesoglea.
- Anthozoa Sea anemones and corals. Have only the polyp stage.
- Hydrozoa Although the freshwater hydra is a much-studied representative, it is not typical of the class.
Bilaterians
All the remaining groups of animals belong in a clade whose members share:
- bilateral symmetry (hence the name); that is, dorsal-ventral and left-right axes
- triploblastic (three tissue layers: ectoderm, mesoderm, endoderm)
- HOX genes in one or more clusters with the genes within a cluster arranged in the same order as the body parts they affect.
The bilaterians contain two clades, the protostomia and the deuterostomia.
Protostomia vs. Deuterostomia
Long before the days of genome analysis, taxonomists were convinced of a fundamental division in the animal kingdom between the protostomes ("first mouth") and the deuterostomes ("second mouth").
| Protostomia | Deuterostomia |
|---|---|
| Blastopore forms future mouth (in most groups). | Blastopore forms future anus. Mouth forms later. |
| Few HOX genes for the posterior | Multiple HOX genes for the posterior |
| Spiral cleavage of Lophotrochozoan embryos | Perpendicular cleavage planes in embryo |
| Early cleavage cells committed; no identical twins | Early cleavage cells totipotent; identical twins possible |
| Coelom arises by splitting of mesoderm | Coelom arises between invaginating mesoderm during gastrulation |
| Lophotrochozoans and Ecdysozoans | Echinoderms, Acorn worms, and Chordates |
Let's first examine the protostomes. The deuterostomes are discussed below.
Lophotrochozoans vs. Ecdysozoans
Genome analysis, especially the analysis of 18S rRNA genes and HOX genes supports a major division of the Protostomia into two superphyla: Lophotrochozoans and Ecdysozoans.
Lophotrochozoans
Their name was created from the names of formerly-separated groups that have now been joined in a single clade on the basis of the similarities of their genomes. They all share a cluster of HOX genes quite different from those found in the ecdysozoans (and deuterostomes). They share similar sequences in their 18S rRNA genes. The clade contains a number of phyla of which we shall examine only 3.
- flatworms (Platyhelminthes),
- annelids (Annelida), and
- mollusks (Mollusca).
Flatworms (Phylum Platyhelminthes)
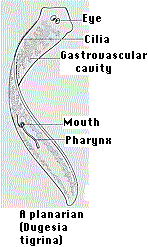
This phylum contains some 20,000 species distributed among three classes. Turbellaria, free-living forms of which the planarian is a commonly-studied example. Planaria share with the other members of the phylum (1) a flat, almost ribbonlike, shape and (2) bilateral symmetry. The bilateral symmetry of planarians is associated with active locomotion by secreting a layer of mucus underneath them and propelling themselves forward with the many cilia on their ventral surface and by swimming and a concentration of sense organs in the head (called cephalization). Planarians feed through a mouth on their ventral surface. It leads to an elaborate gastrovascular cavity. But there is no separate exit so undigested food has to leave by the mouth.
- Trematoda, a group of parasitic
- lung flukes
- liver flukes
- blood flukes (e.g., Schistosoma)
- All of these have at least two different stages in their life cycle, each parasitic in a different host - one of which is usually a snail.
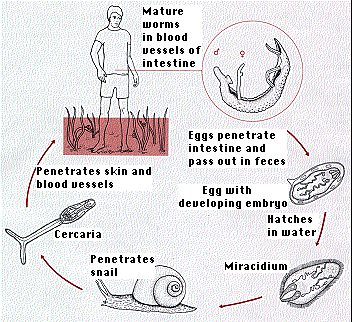
The diagram gives the life cycle of the blood fluke, Schistosoma mansoni. Once within the alternate host, a snail, a single miracidium may produce as many as 200,000 infectious cercariae. Both sexes must infect the human if the cycle is to continue. With the increasing use of irrigation in tropical regions, the incidence of human infection — known as schistosomiasis or bilharzia — is rising alarmingly.
- Cestoda; the parasitic tapeworms. They, too, alternate between an intermediate host (e.g., pig, fish) and a definitive host (e.g., us). The growing popularity of sushi and sashimi made of raw Pacific salmon has caused infections by the fish tapeworm to become more common in the U.S.
Annelids (Phylum Annelida)
Characteristics:
- segmented; that is, their body is made up of repeating units. Although some structures, e.g., the digestive tract, run straight through, others like the excretory organs are repeated in each segment.
- The major nerve trunk runs along the ventral side.
- a large, fluid-filled coelom; It is lined with mesoderm and enables the internal organs to slide easily against one another making for easy locomotion.
There are >15,000 species known. Some examples:
- the common earthworm
- leeches
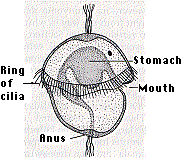
- marine forms such as the clam worm These animals produce a free-swimming trochophore larva (figure), which partly accounts for the name Lophotrochozoan.
Mollusks (Phylum Mollusca)
With over 100,000 living species identified so far, the mollusks must be counted as among the most successful animals on earth today. Most belong to the first 3 of the 6 classes shown here:
- Bivalvia. Two shells encase the body. Includes the clams, mussels, oysters, and scallops.
- Gastropoda. Snails and slugs. Snails have a single shell ("univalves') while slugs have none.
- Cephalopoda. This marine group includes the various species of octopus, squid, as well as the chambered nautilus. A record 28-foot (8.5 m) octopus and 60-foot (18 m) squid make these the largest of all the invertebrates.
- Scaphopoda. Marine, filter-feeding "tooth shells".
- Monoplacophora. Until a live specimen was discovered in 1952, these animals were thought to have been extinct for millions of years. It has a single shell (hence the name) and, unlike the other mollusks, is segmented (as are its relatives the annelids).
- Polyplacophora. The animals in this group, called chitons, have their dorsal surface protected by 8 overlapping plates or "valves".
The trochophore larvae of mollusks is also evidence that they belong in the same clade with the annelids.
Ecdysozoans
All the members of this clade
- grow by periodically molting - shedding their skin or exoskeleton
- share a unique pattern of HOX genes, e.g. Ubx and Abd-B
The clade includes a number of phyla of which we shall examine 2:
- nematodes
- arthropods.
Roundworms (Phylum Nematoda)
Features:
- A one-way digestive tract running from mouth to anus.
- A cavity between the digestive tract and the body wall. It develops from the blastocoel and is called a pseudocoel.
- Some 25,000 species have been identified but this may be less than 10% of the true number.
- Most are free-living; found in soil where they are important decomposers.
- One of these is Caenorhabditis elegans, a model laboratory animal.
- Some are parasitic, including
- hookworms (In 2003 the number of humans infected by hookworms was estimated at 740 million worldwide.)
- pinworms and whipworms
- filarial worms - threadlike worms that are transmitted to the definitive host from an intermediate host causing such human ailments as
- river blindness (Onchocerca volvulus) - acquired from the bite of infected black flies
- elephantiasis (Wuchereria bancrofti) - acquired from infected mosquitoes
- dracunculiasis (Guinea worm disease) (Dracunculus medinensis) - acquired from ingesting water containing infected "water fleas" (Cyclops)
- many parasites of commercially important plants like strawberries and oranges.
- Most are small although one that parasitizes whales reached 30 feet (9 m)!
Arthropods (Phylum Arthropoda)
Some characteristics:
- Incredible diversity. Over a million living species have been identified so far - more than all the other species of living things put together - and this is probably only a fraction of them.
- Live in every possible habitat: fresh water, salt water, soil, even in the most forbidding regions of Antarctica and high mountains.
- A jointed external skeleton made of chitin, a polymer of N-acetylglucosamine (NAG).
- Segmented.
- Pairs of jointed appendages; one pair to a segment - used for locomotion, feeding, sensation, weaponry.
- Bilateral symmetry.
- Main nerve cord runs along the ventral side.
We shall look at four groups (subphyla):
- Crustacea
- Hexapoda (the insects)
- Myriapoda
- Chelicerata
Crustacea
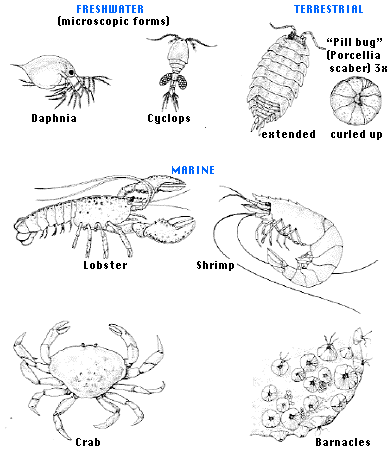
Figure 19.1.10.7 Crustacea
- Head and thorax fused into a cephalothorax.
- At least 40,000 species.
- Most are aquatic, found in both fresh water and in the oceans.
- Includes crayfish, lobsters, barnacles, crabs, shrimp.
Hexapoda - the insects
- Body segments grouped into head, thorax, and abdomen.
- Each of the 3 thoracic segments carries a pair of legs (hence the 6-legged "hexapoda")
- Many have wings, usually 2 pairs (only one pair in flies - diptera).
- Gas exchange through a tracheal system.
- Nitrogenous waste is uric acid thus conserving water.
- Some 950,000 species, and this may be only 10% of the number out there.
- Dominate all habitats except for the oceans.
- Most intensively-studied representative: Drosophila melanogaster.
- Representative colonial insect: the honeybee, Apis mellifera
Myriapoda
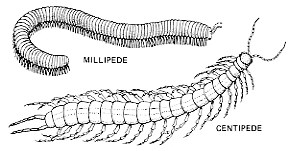
Some 13,000 species of
- centipedes
- millipedes
Neither group has the number of legs their name suggests, although one species of millipede does have 375 pairs.
Chelicerata
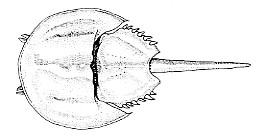
- Anterior segments fused into a cephalothorax.
- The first pair of appendages - the chelicerae - are used for feeding.
- There are no antennae.
- Includes:
- Merostomata. The only member alive today is Limulus, the horseshoe "crab". It has existed in the sea virtually unchanged for 200 million years.
- Arachnids (some 75,000 species)
- 8-legged
- scorpions, mites, ticks, spiders, daddy longlegs.
Evolutionary relationships of the arthropods
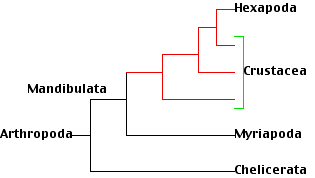
An ever-increasing number of arthropod gene sequences appear to have answered some long-standing questions about the evolutionary relationships of the various arthropod groups. A recent study (Regier, J. C., et al., Nature, 463:1079, 25 February 2010) examined 63 nuclear genes from 75 species of arthropods and concluded that
- the crustacea are paraphyletic; that is, the single common ancestor from which all the animals we call crustaceans are descended was also the ancestor of another group, the insects (Hexapoda). So insects are terrestrial crustaceans!
- All these groups plus the millipedes and centipedes (Myriapoda) make up a clade designated Mandibulata.
- So millipedes and centipedes are more closely related to the crustaceans than to, as once thought, the Chelicerata.
The Deuterostomes
In addition to the features listed above, the deuterostomes have (or had) gill slits. (The echinoderms have lost the gill slits of their ancestors.)
Echinoderms (Phylum Echinodermata)

Characteristics:
- radial symmetry. HOWEVER, their larvae have bilateral symmetry so the echinoderms probably evolved from bilaterally symmetrical ancestors and properly belong in the Bilateria.
- water vascular system. Seawater is taken into a system of canals and is used to extend the many tube feet. These have suckers on their tips and aid the animal in attaching itself to solid surfaces.
- no gill slits
- About 6,000 species — all of them marine.
There are 5 classes of echinoderms:
- Sea lilies (Crinoidea)
- Sea Stars (aka "Starfish") (Asteroidea) The photo (courtesy of Dr. Charles Walcott) shows a sea star that lost an arm and is in the process of regenerating a replacement.
- Brittle stars (Ophiuroidea)
- Sea cucumbers (Holothuroidea)
- Sea urchins and sand dollars (Echinoidea)
Acorn Worms (Phylum Hemichordata)
The members of this small phylum (some 90 species have been identified) are marine forms most of whom live in burrows in ocean sediments. Their closest living relatives are the echinoderms with which they share the clade Ambulacraria. However, they possess a suite of features, both in their anatomy (e.g. gill slits) and their gene expression patterns, suggesting that their ancestors also led to the evolution of the chordates.
Chordates (Phylum Chordata)
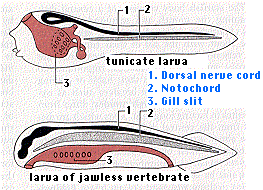
During their embryonic development, all chordates pass through a stage called the pharyngula with these features:
- a dorsal, tubular nerve cord ("1") running from anterior to posterior. At its anterior end, it becomes enlarged to form the brain.
- a flexible, rodlike notochord ("2") that runs dorsal to the digestive tract and provides internal support. In vertebrate chordates, it is replaced by a vertebral column or backbone long before maturity.
- pairs of gill pouches. These lateral outpocketings of the pharynx are matched on the exterior by paired grooves. In aquatic chordates, one or more pairs of gill pouches break through to the exterior grooves, forming gill slits ("3"). These provide an exit for water taken in through the mouth and passed over the gills.
- a tail that extends behind the anus
- The vast majority of chordates have a skull enclosing their brain (Craniata), and all but one of these (the hagfish) convert their notochord into a vertebral column or backbone. These latter are the vertebrates.
Vertebrates also differ from all the other animals by having quadrupled their HOX gene cluster; that is, vertebrates have 4 clusters of HOX genes located on 4 different chromosomes.
Here we shall examine two groups of invertebrate chordates:- Urochordata and
- Cephalochordata
Urochordata
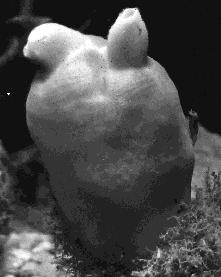
This group (also called Tunicata) includes animals known as ascidians (and commonly called sea squirts). They are
- marine
- sessile animals
- feed by filtering food particles from seawater taken in through one opening, or siphon, and squirted out the other.
The one on the above is Halocynthia, the sea peach (photo courtesy of Ralph Buchsbaum). It is hard to see what makes these animals chordates. The adults have neither notochord nor a dorsal tubular nervous system. However, these animals disperse themselves with free-swimming larvae that have
- dorsal tubular nervous system
- notochord
- gill slits
One of the most common species (Ciona intestinalis) has had its genome sequenced.
- It has a very small genome: ~1.6 x 108 base pairs encoding ~16,000 genes. (Some 20% of these are organized in operons.)
- Its larva is small (with ~2,600 cells) including only
- 36 muscle cells
- 40 notochord cells
- 100 neurons
- These cells (as well as the others) develop along rigid pathways which can be easily observed because the larva is
- transparent.
All these features are shared with C. elegans, but now we are talking about an animal far closer to the evolutionary line that produced us. In fact, with 80% of Ciona's genes having homologs in us, tunicates are probably our closest invertebrate relatives.
Cephalochordata
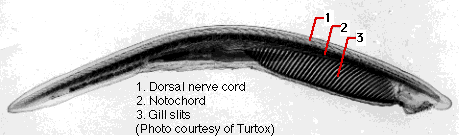
The representative member of this tiny subphylum of so-called lancelets is a small (5 cm), marine, fishlike creature called amphioxus (above). For years its genus name was Amphioxus but that has now been replaced by the name Branchiostoma. Amphioxus retains a dorsal nerve cord, notochord and gill slits throughout its life. There is a small cluster of neurons at the anterior tip of the nerve cord with certain similarities of structure and gene expression to the vertebrate fore-, mid- and hindbrain. Although able to swim, the lancelet spends most of its time partially buried in the sand while it filters microscopic food particles from the water.
Contributors and Attributions
John W. Kimball. This content is distributed under a Creative Commons Attribution 3.0 Unported (CC BY 3.0) license and made possible by funding from The Saylor Foundation.