
19.1.2: Protists
- Page ID
- 5943

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)What are protists?
- They are eukaryotes because they all have a nucleus.
- Most have mitochondria although some have later lost theirs. Mitochondria were derived from aerobic alpha-proteobacteria that once lived within their cells.
- Many have chloroplasts with which they carry on photosynthesis. Chloroplasts were derived from photosynthetic cyanobacteria living within their cells.
- Many are unicellular and all groups (with one exception) contain some unicellular members.
- The name Protista means "the very first", and some of the 80-odd groups of organisms that we classify as protists may well have had long, independent evolutionary histories stretching as far back as 2 billion years. But genome analysis added to other criteria show that others are derived from more complex ancestors; that is, are not "primitive" at all.
- Genome analysis also shows that many of the groups placed in the Protista are not at all closely related to one another; that is, the protists do not represent a single clade.
- So we consider them here as a group more for our convenience than as a reflection of close kinship, and a better title for this page would be "Eukaryotes that are neither Animals, Fungi, nor Plants".
The Euglenozoa
Most Euglenozoa are unicellular. Many swim by means of a single flagellum. They are not encased in a cell wall so they are flexible as well as motile. Euglena is a typical member of the group (which numbers about 1600 species). Because some members of the group (like Euglena) have chloroplasts, these organisms used to be called "Euglenophytes", but in fact they are neither plants ("phytes") nor animals ("zoa"). Rather — like the other organisms on this page — they are the living descendants of some of the very earliest eukaryotes. Trypanosoma brucei, the cause of African sleeping sickness in humans, is a member of the group. The electron micrograph shows T. brucei as it occurs in the salivary gland of the tsetse fly ready to be injected into the mammalian host when the fly bites. The specimen is 12 µm long.
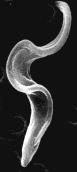
In Latin America, Trypanosoma cruzi, another member of the group, is the cause of Chagas disease in humans.
Ciliates, Sporozoans, and Dinoflagellates: the Alveolates
These three phyla are grouped in a clade called the alveolates because they all have a system of saclike structures ("alveoli") on the inner surface of their plasma membrane as well as close homology in their gene sequences.
Ciliates
The ciliates move by the rhythmic beating of their cilia. Although single-celled, some are large enough to be seen with the naked eye. In fact, the tiny parasitic wasp Megaphragma mymaripenne, with its tens of thousands of cells (4,600 neurons alone), is no larger than Paramecium. They feed by sweeping a stream of particle-laden water through a "mouth" and "gullet" and into a food vacuole. Undigested wastes are discharged at a permanent site. Fresh water ciliates cope with the continuous influx of water from their hypotonic surroundings by pumping it out with one or more contractile vacuoles. Parasitic ciliates, which live in isotonic surroundings, have no contractile vacuole. All of this rightly suggests that although they are unicellular, there is nothing rudimentary about the ciliates. Their single cell is far more elaborate in its organization than any cell out of which multicellular organisms are made. Examples: Paramecium, Stentor, Vorticella, Tetrahymena thermophila.
Sporozoans (Apicomplexa)
The members of this group share an "apical complex" of microtubules at one end of the cell (hence the name that many prefer to the old name of sporozoans). All the members of the phylum are parasites. The genus Plasmodium causes malaria, one of the greatest scourges of humans. There are 4 species that infect humans of which Plasmodium falciparum is the most dangerous. Malaria has probably caused more human deaths than any other infectious disease; even today it is estimated to kill a million people a year in the sub-Saharan Africa. The organism is transmitted from human to human through the bite of mosquitoes of the genus Anopheles.

The diagram shows the Plasmodium life cycle.
- The mosquito bite injects sporozoites into the human host.
- These invade the liver where they develop into merozoites.
- The merozoites invade red blood cells where they reproduce.
- Periodically, they all break out of the red cells together bringing on the chills and fever characteristic of the disease.
- Eventually some merozoites develop into either male or female gametocytes.
- These will die unless they are sucked up by the bite of an anopheline mosquito.
- Once in the stomach of the mosquito, the gametocytes form gametes: sperm and eggs.
- These fuse to form zygotes.
- The zygote invades the stomach wall of the mosquito forming thousands of sporozoites.
- These migrate to the salivary gland, ready to be injected into a new human host.
- Most forms of malaria are chronic. The organisms may coexist with their host for years (but cannot complete their life cycle there).
Toxoplasma gondii is another parasitic member of this group. Plasmodium, Toxoplasma, and some of the other members of this group contain a membrane-enclosed organelle called the apicoplast. They seem to have inherited it from a common ancestor that acquired it by engulfing a chloroplast.
Dinoflagellates
There are about 1000 species of dinoflagellates. Most are unicellular. Most use chlorophylls a and c. Unlike most eukaryotes, they lack histones on their chromosomes and have a simpler form of mitosis. They do have the eukaryotic type ("9 + 2") of flagellum (two of them in fact). Occasionally they reproduce explosively, creating poisonous red tides that may cause extensive kills of marine fish and make filter-feeding marine animals like clams unfit for human consumption.
The Stramenopiles
These organisms belong to a single clade, the stramenopiles (a/k/a heterokonts). There are four members in this group - diatoms, golden algae, brown algae and water molds. The first three members share:
- a yellow-brown pigment (which gives them their color). It is a carotenoid called fucoxanthin.
- chlorophylls a and c
- All four of them (plus a number of other groups not listed) share genes closely-homologous to those in both green and red algae. This suggests that they are all descended from a heterotrophic eukaryotic ancestor that acquired both a green alga and a red alga by a secondary endosymbiosis. (While the water molds no longer are photosynthetic, they still retain both green and red alga genes.)
Diatoms
Diatoms are unicellular. Their cell wall or shell is made of two overlapping halves. These are impregnated with silica and often beautifully ornamented. The photo (courtesy of Turtox) is of Arachnoidiscus ehrenbergi magnified some 400 times. Diatoms are major producers in aquatic environments; that is, they are responsible for as much as 40% of the photosynthesis that occurs in fresh water and in the oceans. They serve as the main base of the food chains in these habitats, supplying calories to heterotrophic protists and small animals. These, in turn, feed larger animals.

Golden Algae (Chrysophyta)
- Most are unicellular.
- Found in fresh water.
- Important producers in some aquatic food chains.
- In low light conditions, may lose their chlorophyll and turn heterotrophic feeding on bacteria and/or diatoms.
- Over 1000 species alive today; many more in the fossil record.
Brown Algae (Phaeophyta)
- The rockweeds and kelps. Some kelps grow as long as 30 meters.
- All are multicellular although without much specialization of cell types.
- Most are found in salt water.
- Used for food in some coastal areas of the world and harvested in the U. S. for fertilizer and as a source of iodine.
Water Molds (Oomycetes)
As their name suggests, water molds were once considered to be fungi. But unlike fungi, the cell wall of water molds is made of cellulose, not chitin. Furthermore, their gene sequences are very different from those of fungi (and most closely related to those of diatoms, golden and brown algae).
Some notable water molds:
- Some species (e.g., Saprolegnia, Achyla) are parasites of fishes and can be a serious problem in fish hatcheries.
- Downy mildews damage grapes and other crops.
- Phytophthora infestans, the cause of the "late blight" of potatoes. In 1845 and again in 1846, it was responsible for the almost total destruction of the potato crop in Ireland. This led to the great Irish famine of 1845–1860. During this period, approximately 1 million people starved to death and many more emigrated to the New World. By the end of the period, death and emigration had reduced the population of Ireland from 9 million to 4 million.
- Phytophthora ramorum, which is currently killing several species of oaks in California.
Red Algae
The red algae are almost exclusively marine. Some are unicellular but most are multicellular. Approximately 6000 species have been identified. They are photosynthetic using chlorophyll a. Their closest relatives are the green algae and land plants. Like the cyanobacteria, they use as antenna pigments - phycoerythrin (which makes them red) and phycocyanin. They do not have the eukaryotic "9+2" flagellum. Some are used as food in coastal regions of Asia. Agar, the base for culturing bacteria and other microorganisms, is extracted from a red alga.
Slime Molds (Mycetozoa)
Cellular Slime Molds
The organisms in this group have a complex life cycle during the course of which they go through unicellular, multicellular, funguslike (form spores) and protozoanlike (amoeboid) stages. Thousands of individual amoebalike cells aggregate into a slimy mass - each cell retaining its identity (unlike plasmodial slime molds). The aggregating cells are attracted to each other by the cyclic AMP (cAMP) that they release.
With the exception of one species that causes powdery scab on potatoes, these organisms are of little economic importance. However, their combination of traits makes them of great scientific interest. Molecular phylogenies place them in the same clade as animals (metazoa) and fungi.
Plasmodial (Acellular) Slime Molds (Myxomycetes)


At one stage in their life cycle, these organisms consist of a spreading, slimy, multinucleate mass called a plasmodium that moves slowly over its substrate (e.g., a rotting log) engulfing food and growing as it does so. Eventually, the plasmodium develops stalks that produce and release spores. If the spores land in a suitable location, they germinate forming single cells that move by both flagella and pseudopodia. These fuse in pairs and start forming a new plasmodium.
The left photo (courtesy of Prof. I. K. Ross) shows the plasmodial stage of Stemonitis just before it formed sporangia. The right photo (courtesy of Turtox) shows the fully developed sporangia of Stemonitis.
Physarum polycephalum, another member of this group, is the subject of many laboratory studies.
Protists without typical mitochondria
There are several groups of protists that were long thought to have no mitochondria. However, most (perhaps all) had them in the past. Today, only remnants of their ancestor's mitochondria - called mitosomes remain.
Some examples are:
- Microsporidia
- All are unicellular obligate intracellular parasites.
- Many are pathogenic in insects (one is even marketed commercially as a biocontrol agent).
- Some contaminate drinking water supplies and can cause gastrointestinal upsets in humans. Microsporidia, such as Encephalitozoon cuniculi, are a common cause of diarrhea in AIDS patients. Encephalitozoon cuniculi has a tiny genome with only 1,997 protein-encoding genes — fewer than many bacteria (e.g., E. coli has 4,290). Obliged to live within the cells of its host, it has lost the genes for many important functions (e.g., the citric acid cycle) depending instead on its host.
- Fungi are their closest relatives.
- Entamoeba histolytica.
- Causes amebic dysentery, the third most common parasitic disease of humans (after malaria and schistosomiasis). Its closest relatives are the slime molds.
- Giardia intestinalis (also known as Giardia lamblia)
- Frequently encountered in public water supplies contaminated by animal feces. Causes diarrhea in humans. Avoids the host immune response by periodically changing its surface protein coat.
Choanoflagellates

These are single-celled (e.g., Monosiga), aquatic (both fresh water and marine) protists that have a single flagellum surrounded by a collar ("choano" = collar) of microvilli. Some (e.g., Proterospongia) form simple colonies during part of their life. The flagellum is used for swimming and also beats bacteria-containing water through the collar for feeding.
Sponges also use collar cells to filter food from the water. Not only does this suggest a close relationship between the two groups, but other evidence indicates that choanoflagellates are the closest protistan relatives of all animals (metazoa). Although single cells, they express genes for several proteins that are essential to cell-cell interactions in metazoans, such as
- cadherins (attach cells to each other)
- tyrosine kinases (used in many examples of cell-cell signaling)
What function these proteins have in the choanoflagellates is unknown.