
16.2F: Tropisms
- Page ID
- 5787

A tropism is a growth movement whose direction is determined by the direction from which the stimulus strikes the plant.
- Positive = the plant, or a part of it, grows in the direction from which the stimulus originates.
- Negative = growth away from the stimulus.
Plants respond to:
- Light = phototropism
- Stems are positively phototropic.
- Roots are negatively phototropic.
- Gravity = gravitropism
- Stems are negatively gravitropic while
- roots are positively gravitropic.
The adaptive value of these tropisms is clear.
- Roots growing down and/or away from light are more likely to find the soil, water, and minerals they need.
- Stems growing up and toward the light will be able to expose their leaves so that photosynthesis can occur.
The Mechanism of Phototropism
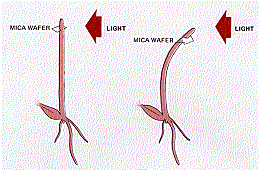
The Darwin Experiments

If they placed an opaque cover over the tip, phototropism failed to occur even though the rest of the coleoptile was illuminated from one side. However, when they buried the plant in fine black sand so that only its tip was exposed, there was no interference with the tropism — the buried coleoptile bent in the direction of the light.
From these experiments, it seemed clear that
- the stimulus (light) was detected at one location (the tip)
- the response (bending) was carried out at another (the region of elongation)
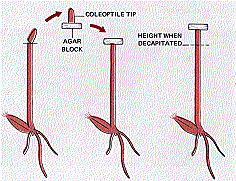
The Danish plant physiologist Boysen-Jensen showed (in 1913) that the signal was a chemical passing down from the tip of the coleoptile.
He
- cut off the tip of the coleoptile
- covered the stump with a layer of gelatin
- replaced the tip.
The graph (based on the work of K. V. Thimann) shows the effect of auxin concentration on root and stem growth. The difference between the behavior of roots and stems lies in the difference in the sensitivity of their cells to auxin. Auxin concentrations high enough to stimulate stem growth inhibit root growth.
Possible Mechanism of Gravitropism in Roots
When a root is placed on its side,
- Statoliths (organelles containing starch grains) settle by gravity to the bottom of cells in the root tip.
- This causes PIN proteins to redistribute to the underside of the cell where they pump auxin out of the cell.
- The auxin then accumulates along the under side of the root.
- This INHIBITS root cell elongation (see graph).
- So the cells at the top surface of the root elongate, causing the root to grow down.