8.3: Meiotic Recombination
- Page ID
- 359

A diploid organism has two copies of each chromosome. If it has four chromosomes, there are two pairs, A and A’ and B and B’, not four different chromosomes A, B, C and D. One copy of each chromosome came from its father (e.g. A and B) and one copy of each came from its mother (e.g. A’ and B’). Meiosis is the process of reductive division whereby a diploid organism generates haploid germ cells (in this case, with two chromosomes), and each germ cell has a single copy of each chromosome. In this example, meiosis does not generate germ cells with A and A’ or B and B’, rather it produces cells with A and B, or A and B’, or A’ and B, or A’ and B’. The homologous chromosomes, each consisting of two sister chromatids, are paired during the first phase of meiosis, e.g., A with A’ and B with B’ (Figure 8.3; see also Figs. 1.3 and 1.4). Then the homologous chromosomes are moved to separate cells at the end of the first phase, insuring that the two homologs do not stay together during reductive division in the second phase of meiosis. Thus each germ cell receives the haploid complement of the genetic material, i.e. one copy of each chromosome. The combination of two haploid sets of chromosomes during fertilization restores the diploid state, and the cycle can resume. Failure to distribute one copy of each chromosome to each germ cell has severe consequences. Absence of one copy of a chromosome in an otherwise diploid zygote is likely fatal. Having an extra copy of a chromosome (trisomy) also causes problems. In humans, trisomy for chromosomes 15 or 18 results in perinatal death and trisomy 21 leads to developmental defects known as Down’s syndrome.
Exercise 8.2
If this diploid organism with chromosomes A, A’, B and B’ underwent meiosis without homologous pairing and separation of the homologs to different cells, what fraction of the resulting haploid cells would have an A-type chromosome (A or A’) and a B-type chromosome (B or B’)?
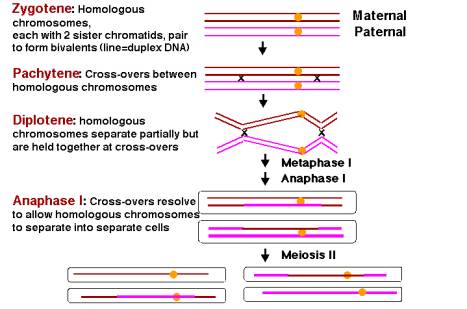
The ability of homologous chromosomes to be paired during the first phase of meiosis is fundamental to the success of this process, which maintains a correct haploid set of chromosomes in the germ cell. Recombination is an integral part of the pairing of homologous chromosomes. It occurs between non-sister chromatids during the pachytene stage of meiosis I (the first stage of meiosis) and possibly before, when the homologous chromosomes are aligned in zygotene (Figure 8.3). The crossovers of recombination are visible in the diplotene phase. During this phase, the homologous chromosomes partially separate, but they are still held together at joints called chiasmata; these are likely the actual crossovers between chromatids of homologous chromosomes. The chiasmata are progressively broken as meiosis I is completed, corresponding to resolution of the recombination intermediates. During anaphase and telophase of meiosis I, each homologous chromosome moves to a different cell, i.e. A and A’ in different cells, B and B’ in different cells in our example. Thus recombinations occur in every meiosis, resulting in at least one exchange between pairs of homologous chromosomes per meiosis.
Recent genetic evidence demonstrates that recombination is required for homologous pairing of chromosomes during meiosis. Genetic screens have revealed mutants of yeast and Drosophila that block pairing of homologous chromosomes. These are also defective in recombination. Likewise, mutants defective in some aspects of recombination are also defective in pairing. Indeed, the process of synapsis (or pairing) between homologous chromosomes in zygotene, crossing over between homologs in pachytene, and resolution of the crossovers in the latter phases of meiosis I (diakinesis, metaphase I, and anaphase I) correspond to the synapsis, formation of a recombinant joint and resolution that mark the progression of recombination, as will be explained below.