
17.7: 17.7 Signal Transduction
- Page ID
- 24559

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)When hydrophobic chemical effector molecules such as steroid hormones reach a target cell they can cross the hydrophobic membrane and bind to an intracellular receptor to initiate a response. When large effector molecules (e.g., protein hormones) or highly polar hormones (e.g., adrenalin) reach a target cell, they can’t cross the cell membrane. Instead, they bind to transmembrane protein receptors on cell surfaces. A conformational change initiated on the extracellular domain of the receptor induces further allosteric change on the cytoplasmic domain of the receptor. A sequential series of molecular events then converts information delivered by the external effector into intracellular information, a process called signal transduction. A general outline of signal transduction events is illustrated below.
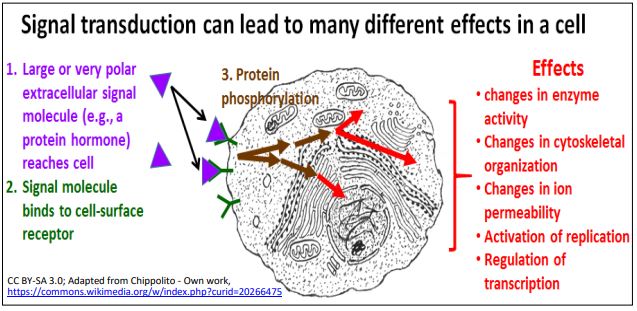
Many effects of signal transduction are mediated by a sequence, or cascade of protein phosphorylations catalyzed by protein kinases inside the cell. Here we will consider G Protein-linked and enzyme-linked receptors.
315 Introduction to Signal Transduction
A. G-Protein Mediated Signal Transduction by PKA (Protein Kinase A)
GTP-binding proteins (G-Proteins) transduce extracellular signals by inducing production of second messenger molecules in the cells. When hormones or other effector (signal) molecules bind to their membrane receptors, an allosteric change on the cytoplasmic domain of the receptor increases the affinity of the cytoplasmic domain the receptor for G proteins on the inner plasma membrane surface. G proteins are trimers consisting of \(\alpha \), \(\beta \) and \(\gamma \) subunits, embedded in the cytoplasmic surface of responsive cell membranes. G-protein-mediated signal transduction is illustrated in the seven steps shown on the next page.
The receptor changes shape upon binding its effector signal molecule (steps 1, 2). In this conformation, the receptor recognizes and binds to the G-protein trimer on the cytoplasmic surface of the plasma membrane (step 3). Upon binding of the trimer to the receptor, GTP displaces GDP on the \(\alpha \) subunit of the G-protein (step 4).
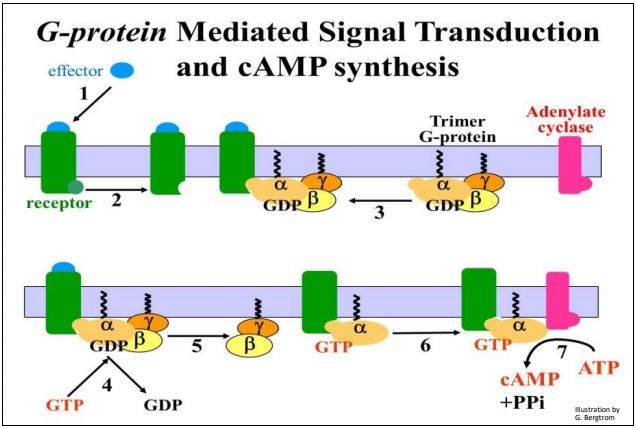
After a conformational change, the \(\alpha \) subunit dissociates from the \(\beta \) and \(\gamma \) subunits (step 5). In this illustration, the GTP-\(\alpha \) subunit can now bind to a transmembrane enzyme, adenylate cyclase (step 6). Finally, the initial extracellular chemical signal is transduced to an intracellular response involving second messenger molecules (step 7). In this case, the second messenger is cAMP. The well-known fight-or-flight response to adrenaline in liver cells of higher animals is a good example of a cAMPmediated cellular response. After adrenalin binds to its receptors, G-proteins in turn bind to the cytoplasmic side of the receptor, which then binds to adenylate cyclase. cAMP binds to and activates protein kinase A (PKA), setting off the amplification cascade response. Some details of a G-protein mediated signal amplification cascade are detailed in the illustration on the next page.
After activation of adenylate cyclase (steps 1 and 2 in the drawing), cAMP is synthesized and binds to two of the four subunits of an inactive PKA (step 3). A conformational change dissociates the tetramer into two cAMP-bound inert subunits and two active PKA subunits (step 4). Each active PKA enzyme catalyzes phosphorylation and activation of an enzyme called phosphorylase kinase (step 5).

In step 6, phosphorylase kinase catalyzes glycogen phosphorylase phosphorylation. Finally, at the end of the phosphorylation cascade, the now active glycogen phosphorylase catalyzes the hydrolysis glycogen to glucose-1-phosphate (step 7). This results in a rapid retrieval free glucose from liver cells into the circulation. Remind yourself of how this works by reviewing the conversion of glucose-1 phosphate (G-1-P) to G-6-P in glycolysis and its fate in gluconeogenesis. Of course, the increase in circulating glucose provides the energy for the fight-or-flight decision.
317 G-Protein Activation of Protein Kinase A and a Fight-or-Flight Response
In addition to activating enzymes that break down glycogen, cAMP-activated PKA mediates cellular responses to different effectors resulting in a phosphorylation cascade leading to
- Activation of enzymes catalyzing glycogen synthesis.
- Activation of lipases that hydrolyze fatty acids from triglycerides.
- Microtubule assembly.
- Microtubule disassembly.
- Mitogenic effects (activation of enzymes of replication).
- Activation of transcription factors increasing/decreasing gene expression.
Of course, when the cellular response is no longer needed by the organism, it must stop producing the signal molecules (hormone or other effector). As their levels drop, effector molecules dissociate from their receptors and the response stops. This is all possible because binding of signals to their receptors is freely reversible! This is animated for G-protein based signal transduction in the link below.
B. Signal Transduction using PKC
Many responses involving G-proteins begin by activating the integral membrane adenylate cyclase. A different G-protein-mediated signaling pathway generates other second messengers. Protein kinase C (PKC) plays a major roles in the activating these other second messengers and subsequent phosphorylation cascades in which the activation of just a few enzyme molecules in the cell results in the activation of many more enzymes. Like PKA, PKC-mediated signal transduction also amplifies the cell’s first molecular response to the effector. The role of G-proteins is similar for PKA and PKC signal transduction. Responses can include diverse effects in different cells…, or even in the same cells using different effector signals. But, PKC and PKA signal transduction differ in that PKC activation requires an additional step, as well as the generation of two intracellular messenger molecules. The events leading to the activation of PKC are illustrated below.
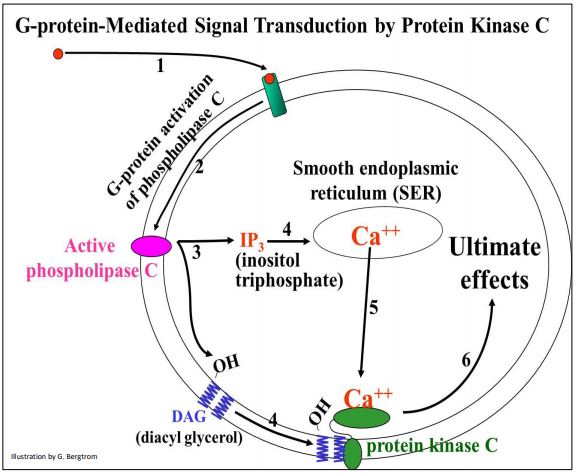
Here are details of the steps leading to PKC activation. An effector signal molecule binds to its receptor, activating an integral membrane phospholipase C enzyme. Phospholipase C catalyzes formation of cytosolic inositol triphosphate (IP3) and membrane bound diacyl glycerol (DAG), two of those other intracellular second messenger molecules. IP3 interacts with receptors on smooth endoplasmic reticulum, causing the release of sequestered Ca++ ions into the cytoplasm. Finally, Ca2+ ions and DAG activate Protein Kinase C (PKC) that then initiates a phosphorylation amplification cascade leading cell-specific responses.
318 G-Protein Activation of Protein Kinase C and Phospholipase C
Protein Kinase C mediated effects include:
- Neurotransmitter release.
- Hormone (growth hormone, leutinizing hormone, testosterone) secretion leading to cell growth, division and differentiation.
- Glycogen hydrolysis, fat synthesis.
Additionaly independent phospholipase C effects include:
- Liver glycogen breakdown.
- Pancreatic amylase secretion.
- Platelet aggregation.
PKA and PKC are serine-threonine kinases that they place phosphates on serine or threonine in target polypeptides. Let’s consider tyrosine kinases next.
C. Receptor Tyrosine Kinase-Mediated Signal Transduction
The intracellular activity of these receptors is in the cytoplasmic domain of the receptor itself. When bound to its effector, receptor-kinase catalyzes phosphorylation of specific tyrosine amino acids in target proteins. While studying the action of nerve growth factor (NGF) and epidermal growth factor (EGF) in stimulating growth and differentiation of nerve and skin, Stanley Cohen and Rita Levi-Montalcini discovered the EGF receptor, the first enzyme-linked tyrosine kinase…, and won the 1986 Nobel Prize in Physiology or Medicine! Watch the animation of receptor kinase signal transduction at the link below (a description is provided in the next few paragraphs).
319 Receptor Kinase Signal Transduction
Monomer membrane receptor kinases dimerize when they bind effector ligands, at which point sulfhydryl group-containing SH2 proteins bind to each monomer. This activates the kinase domain of the receptor. After multiple cross-phosphorylations of the receptor monomers, the SH2 proteins fall away allowing the receptors to interact with other cytoplasmic proteins to continue the response pathway. The characteristic response to EGF and NGF signaling is cellular proliferation. Not surprisingly, mutations correlated with cancer cells often lie in signaling pathways leading to cell proliferation (growth and division). Cancer-causing genes, or oncogenes, were actually first discovered in viruses, but J. Michael Bishop and Harold Varmus won the 1964 Nobel Prize in Physiology or Medicine for showing that cells were actually the origin of a chicken retrovirus (the Rous Sarcoma Virus). Oncogenes turn out to be mutations of genes for proteins in mitogenic signal transduction pathways. Under normal circumstances, mitogenic chemical signals (like EGF) bind to their receptors and induce target cells to begin dividing.
The Ras protein-mediated activation of a phosphorylation cascade leading to the MAP (mitogen-activated protein) kinase is an example of such a signal transduction pathway, one with a central role in many receptor kinase signaling pathways. The Ras gene was one of those originally discovered as an oncogene whose mutation leads to uncontrolled cell division, i.e., cancer. Ras gene/protein activity may in fact be responsible for up to 30% of all cancers!
320 The RAS Oncogene, its Normal Mitogenic Effects and Cancer
MAP kinase phosphorylates transcription factors and other nuclear proteins that affect gene activity leading to cell proliferation and differentiation, as shown below.

D. Signal Transduction in Evolution
We saw that signal transduction typically takes a few signal molecules interacting with a few cell surface receptors to amplify a response in a cascade of enzymatic reactions, typically phosphorylations, to activate (or inactivate) target proteins. Amplification cascades can take a single effector-receptor interaction and magnify its effect in the cell by orders of magnitude, making the signaling systems rapid and highly efficient. The range of cellular and systemic (organismic) responses to the same chemical signal is broad and complex. Different cell types can have receptors for the same effector, but respond differently. For example, adrenalin targets cells of the liver and blood vessels among others, with different effects in each. As it happens, adrenaline is also a neurotransmitter. Apparently, as organisms evolved, they became more complex in response to environmental imperatives, adapting by coopting already existing signaling systems in the service of new pathways. Just as the same signal transduction event can lead to different pathways of response in different cells, evolution has allowed different signal transduction pathways to engage in crosstalk. This is when two different signal transduction pathways intersect in the same cells. In one example, the cAMP produced at the front end of the PKA signaling pathway can activate (or under the right circumstances, inhibit) enzymes in the MAP kinase pathway. These effects result in changes in the levels of active or inactive transcription factors and can therefore modulate the expression of a gene using two (or more) signals. We are only beginning to understand what looks less like a linear.