
17.4: Endocytosis and Exocytosis
- Page ID
- 16523

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Endocytosis is a mechanism for internalizing large extracellular molecules (e.g., proteins), insoluble particles, or even microorganisms. The three main types of exocytosis are phagocytosis, pinocytosis and receptor-mediated endocytosis. Pinocytosis is non-specific. Phagocytosis targets large structures (e.g., bacteria, food particles…) and is not particularly specific. As its name suggests, receptor-mediated endocytosis is specific for substances recognized by a cell-surface receptor. Exocytosis is typically the secretion of large molecules. These could be proteins and glycoproteins like digestive enzymes and many peptide/polypeptide hormones, each of which must exit the cell to either the extracellular fluid or circulation. Exocytotic pathways also deliver membrane proteins made in cells to the cell surface.
A. Endocytosis
Different forms of endocytosis are illustrated on the next page.
1. Phagocytosis (above left): phagocytes extend pseudopodia by membrane evagination. The pseudopodia of amoeba (and amoeboid cells generally) engulf particles of food that end up in digestive vesicles (phagosomes) inside the cytosol. Phagocytes are a class of white blood cells that are part of our immune system. They engulf foreign particles that must be eliminated from the body. A lysosome fuses with the phagosome, after which stored hydrolytic enzyme are activated. The result is the digestion of the engulfed particles. Phagocytosis begins upon contact between the outer cell surface and those particles. The main kinds of endocytosis are phagocytosis, pinocytosis and receptor-mediated endocytosis, shown below.
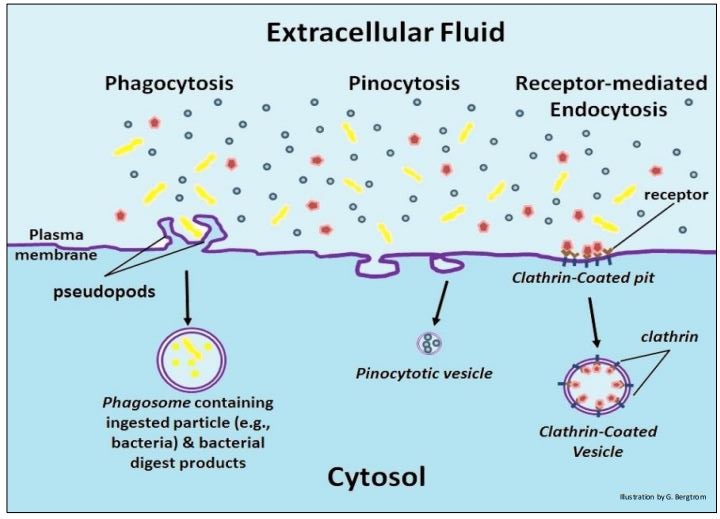
2. Pinocytosis (above center): pinocytosis is a non-specific, more or less constant pinching off of small vesicles that engulf extracellular fluid containing solutes; they are too small to include significant particulates.
3. Receptor-mediated endocytosis (above right): this kind of endocytosis relies on the affinity of receptors for specific extracellular substances. Upon binding their ligands, the receptors aggregate in differentiated regions of cell membrane called coated pits. The coated pits then invaginate and pinch off, forming a coated vesicle, thereby bringing their extracellular contents into the cell. After the coated vesicles deliver their contents to their cellular destinations, the vesicle membranes are recycled to the plasma membrane. Receptor-mediated endocytosis is perhaps the best understood mechanism for bringing larger substances into cells. The drawings below are taken from a series of electron micrographs that illustrates the invagination of coated pits to form clathrin-coated vesicles. The receptor and coat proteins are clearly visible as larger structures on the inner surfaces of the pits and on the outer surfaces of the clathrin-coated vesicles.

Watch fluorescently labeled proteins enter cells by receptor-mediated endocytosis live by following the bright spots in the video loop at Receptor-mediated endocytosis. Clathrin, a large protein, is the principal protein on the surface of the invaginated coated pit. Clathrin is linked to specific integral membrane proteins via adaptor protein 1 (AP1). AP1 recruits specific cargo proteins to bring into the cell when the coated pits invaginate. Some details of receptor-mediated endocytosis are illustrated below.

In the illustration, substances to be internalized have bound to their cell membrane receptors. The receptors then cluster to form a coated pit. Assisted by the protein dynamin (a GTPase), the coated pits invaginate. The final pinch-off of a coated vesicle requires GTP hydrolysis (not shown).
Once internalized, the coated vesicles lose their clathrin and associated adaptor protein coat. The uncoated vesicle fuses with an early endosome to form a sorting vesicle (i.e., late endosome). Sorting vesicles separate imported content from the receptors that are recycled to the membrane. In the vesicle that remains, now a lysosome, digestive enzymes catalyze hydrolysis of the vesicle contents. The digest products are then released for cellular use.
A well-known example of receptor-mediated endocytosis is the uptake of cholesterol bound to low density lipoprotein (LDL), a complex of phospholipid, protein and cholesterol illustrated below.

A single LDL complex carries as many as 15,000 molecules of cholesterol. LDL, sometimes called “bad cholesterol”, is not good for you at high levels. On the other hand, high-density lipoprotein (HDL) is “good cholesterol”. As one gets older, it is important to monitor one’s HDL/LDL ratio; the higher it is the better!
B. Exocytosis
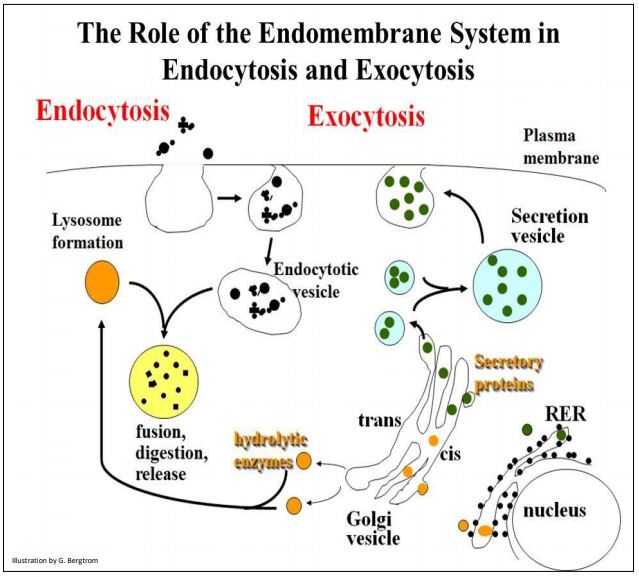
Maintaining cell size or volume seems to be a built-in component of the machinery of receptor-mediated endocytosis that balances endocytosis with membrane recycling. However, exocytosis is also necessary to restore plasma membrane internalized by pinocytosis and phagocytosis, and for eliminating cellular waste products. Exocytosis is also the end-point of a complex process of packaging proteins destined for secretion or for insertion into the membrane themselves. The pathways of exocytosis and endocytosis share common features, as illustrated on the next page.
Note that the formation of both lysosomes and secretion vesicles begins in the rough endoplasmic reticulum, followed by passage and maturation through Golgi vesicles. While endocytotic vesicles and secretion vesicles form in ‘opposite directions’, they both share common structural features with the plasma membrane, from which they are derived and with which they fuse (respectively).
The table on the next page lists some representative proteins packaged for secretion or destined to live in cell membranes.
| Hormones | Immune System Proteins | Neurotransmitters | Other |
| Insulin | IgG (immunoglobulin G, a class of circulating antibodies) | acetylcholine | EGF (Epidermal growth factor) |
| Growth Hormone | IgM and other cell membrane antibodies | NGF (neural growth factor) | |
| FSH (follicle stimulating hormone) | MHC (major histocompatability complex) proteins on cell surfaces | dopamine, adrenaline, noradrenaline & other monoamines | Fibrinogen (& other blood clotting factors) |
| Oxytocin | serotonin | Fibronectin (and other extracellular matrix proteins) | |
| Prolactin | some amino acids (glutamate, aspartate, glycine) | Plant cell wall components | |
| ACTH (adrenocorticotropic hormone) | Trypsin, pepsin, et al. (digestive enzymes of the gut) |
As we have seen, many secretory and membrane proteins are glycoproteins, to which sugars are covalently attached starting in the rough endoplasmic reticulum. As we have also seen, their glycosylation begins in the RER. Check the link below to see the process again.
291 The Path to Sugar Coated Cells
Individual cells often produce more than a few packaged proteins at the same time, requiring the sorting of each protein to its correct destination – extracellular fluids, lysosomes, peroxisomes and other ‘microbodies’, and of course, membranes themselves. Next we consider how cells target proteins to their different intracellular and extracellular destinations.