
17.3: Ligand and Voltage Gated Channels in Neurotransmission
- Page ID
- 16522

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)A. Measuring Ion Flow and Membrane Potential
When neurotransmitters bind to their receptors, ion channels in responding neuron or muscle cells open. The resulting influx of Na+ ions disrupts the resting potential of the target cell. The effect is only transient if the membrane potential remains negative. However, if enough Na+ ions enter the cell, the membrane becomes depolarized. If the cell experiences hyperpolarization, a localized reversal of normal membrane polarity (say from –70 mV to +65mV or more) will generate an action potential. This action potential will travel like a current along the neural or muscle cell membrane, eventually triggering a physiological response, e.g., the excitation of the next nerve cell in a neuronal pathway or contraction of the muscle cell. The patch-clamp device detects specific ion flow and any the resulting change in potential difference across the membrane. Principles of patch-clamp measurement are illustrated below.
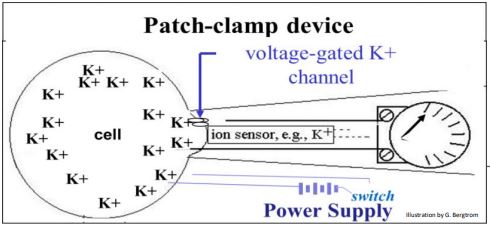
In the example above, closing the switch on the power supply sends an electrical charge to the cell, opening up voltage-gated ion channel. In this case, a potassium sensor in the device detects the flow of K+ ions through the channel and out of the cell. At the same time, a volt meter registers the resulting change in membrane potential.
297 A Patch Clamp Device Can Record Membrane Potential and Ion Flow
298 Patch Clamp Measures Resting Potential and Depolarization
In addition to voltage-gated ion channels, the patch clamp device can measure ion flow through ligand-gated ion channels and mechanically-gated ion channels.
The former channels are receptor-ion gates that open when they bind an effector molecule. Mechanically-gated ion channels detect physical pressure or stress that result in a local membrane deformation, opening the channel.
300 Types of Gated Ion Channels-Illustrated
Finally, cells maintain a high intracellular concentration of K+ ions, causing K+ ions to slowly leak from the cell, a phenomenon detectable by a patch-clamp. The presence of negative ions (Clions, organic ions) inside a cell limits the leakage. This creates the electronegative interior of a cell relative to outside the cell, i.e., the resting potential across its plasma membrane. The patch-clamp technique has been used to correlate the flow of ions and changes in membrane potential when a neuron fires, causing an action potential in a responding cell.
Such a correlation is described on the next page. In the illustration, follow the opening and closing of ion channels and the flow of ions. An action potential (in fact any shift from resting potential) results from facilitated diffusion of specific ions into or out of the cell through gated ion channels (green, above) that must open and close in sequence. The behavior of two different voltage-gated ion channels are illustrated in the graph. Electrical stimulation opens Na+ channels. Na+ ions rush into the cell, reducing the membrane potential from the resting state to zero, or even making the cytoplasm more positive than the extracellular fluid. If the reversal in polarity is high enough, a voltagegated K+ opens and potassium ions rush into the cell, restoring the resting potential of the cell.
A cell can continue to respond to stimuli with action potentials for as long as there is sufficient Na+ outside the cell and K+ inside the cell. While active transport of Na+ and K+ is not required to re-establish the resting potential, it will eventually be necessary to restore the balance of the two ions in the cell. If a nerve or muscle cell fires several times (or even if it just leaks ions), the [K+] inside the cell and the [Na+] outside the cell would drop to a point where the cell cannot generate an action potential when stimulated. Ultimately, it is the role of ATP-dependent Na+/K+ pumps to restore the appropriate Na+:K + balance across the responding cell membrane. As we have seen, each cycle of pumping exchanges 3 Na+ ions from the intracellular space for 2 K+ ions from the extracellular space. The pump has two effects:
- It restores Na+ concentrations in the extracellular space relative to the cytoplasm.
- It restores K+ concentrations in the cytoplasm relative to the extracellular space.
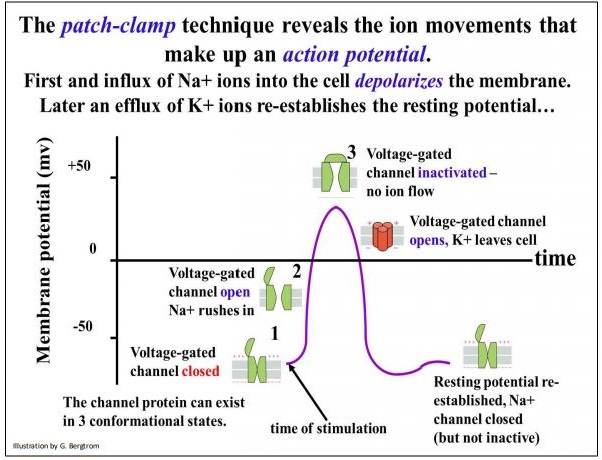
301 Gated Ion Channels Open and Close in Order During an Action Potential
Together with the higher negative ion concentrations in the cytosol, the unequal exchange of Na+ for K+ ions maintains the resting potential of the cell over the long term and ensures that nerve and muscle cells remain excitable. Next, we will take a closer look at the role of both ligand-gated and voltage-gated ion channels in neurotransmission.
B. Ion Channels in Neurotransmission
Action potentials result in an orderly, sequential opening and closing of voltage- and ligand-gated channels along the neuronal axon. In the link below, you can see the sequential cycling of voltage-gated channels that propagates a localized action potential (membrane depolarization) along an axon towards a synapse.
302 Propogating an Action Potential Along an Axon
When a propagated depolarization reaches a synapse, gated ion channels either open or close in the neuron and the responding cell. The cooperation of voltage- and ligand-gated channels at a neuromuscular junction is illustrated below.
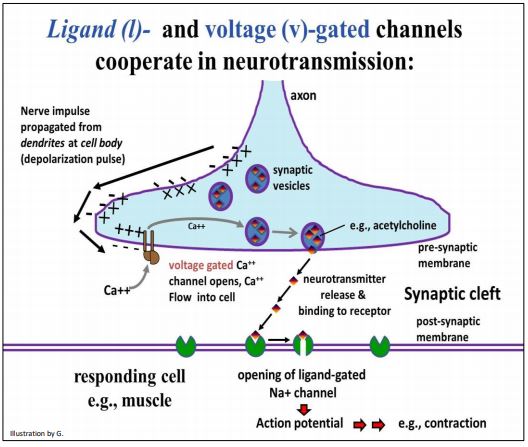
As you can see from the illustration, after a neuron fires, an electrical impulse (a moving region of hyperpolarization) travels down the axon to the nerve ending. At the nerve ending, the traveling charge difference (electrical potential) across the cell membrane stimulates a Ca++ -specific voltage-gated channel to open. Ca++ ions then flow into the cell because they are at higher concentrations in the synaptic cleft than in the cytoplasm.
The Ca2+ ions in the cell cause synaptic vesicles to fuse with the membrane at the nerve ending, releasing neurotransmitters into the synaptic cleft. Then, the neurotransmitters bind to a receptor on the responding cell plasma membrane. This receptor is a ligand-gated channel (also called a chemically-gated channel). Upon binding of the neurotransmitter ligand, the channel opens. The rapid diffusion of Na+ ions into the cell creates an action potential that leads to the cellular response, in this case, muscle contraction. We have already seen that K+ channels participate in restoring membrane potential after an action potential, and the role of the sodium/potassium pump in restoring the cellular Na+/K+ balance.
303 The Role of Gated Ion Channels at Neuromuscular Junction