
17.5: Directing the Traffic of Proteins in Cells
- Page ID
- 16524

Each polypeptide protein translated by ribosomes from a sequence of bases in an mRNA has a specific functional location, either in the cytoplasm, on cellular membranes, inside organelles or in extracellular fluids. In this section we consider the movement and sorting of proteins in the endomembrane system as well as the transport of proteins into and out of organelles
A. Packaging Proteins in the RER
All protein synthesis begins in the same way, with the formation of an initiation complex and subsequent elongation cycles peptide bond formation and carboxylterminal amino acid addition. However, secretory proteins and those destined for lysosomes, peroxisomes or other microbodies, complete elongation directly into cisternae, or spaces enclosed by the rough endoplasmic reticulum (RER). It is possible to isolate and purify proteins secreted by cultured cells. A good model system for studying secretory protein synthesis turn out to be mouse myeloma cells.
Mouse myeloma cells were isolated and grown in culture, where they synthesize an IgG light chain, a polypeptide that is part of a mouse immunoglobulin molecule. Immunoglobulins are assembled from light and heavy chain polypeptides and secreted into the circulation. There they serve as circulating antibodies of the vertebrate immune system. Mouse myeloma cells are cancer cells that have lost the ability to make the heavy chain polypeptides. Instead, they secrete mostly the IgG light chain, making it easy to purify it from the cell culture medium. An early experiment revealed that secreted polypeptides made in an in vitro translation system are larger (longer) than the same polypeptides isolated from secretion fluids. This experiment is summarized on the next page.
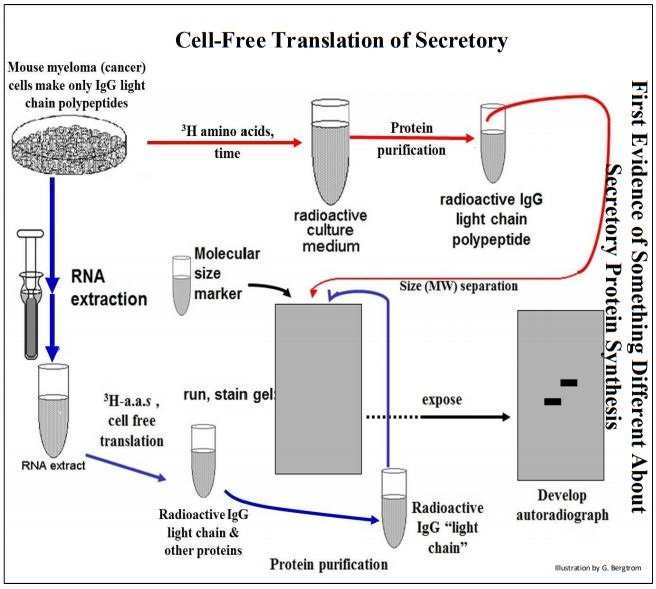
In one part of the experiment described above, myeloma cells were grown in the presence of radioactive amino acids. The resulting radioactive IgG light chain polypeptides were isolated (follow the red arrows). mRNA separately extracted from another batch of the myeloma cells was added to a cell-free translation system containing radioactive amino acids. The radioactive polypeptide synthesized in vivo and in vitro were separated on electrophoretic gels and autoradiographed (follow the blue arrows, above).
From the autoradiograph, the mature, secreted polypeptides made in vivo had migrated faster on the gel than had those translated in vitro. The cell-free translation product was indeed, larger than the mature secreted polypeptide. To explain these results, Gunther Blobel and colleagues suggested the Signal Hypothesis, according to which secretory protein genes encode extra amino acids as a short amino-terminal signal peptide that directs a growing secretory polypeptide to the RER. To explain the smaller (i.e., shorter) length of the mature, secreted polypeptide, they further proposed that the signal peptide is only a temporary ‘traffic’ signal, removed by an RER-associated enzyme as the polypeptide crossed the RER membrane into the cisternal space.
304 Formulating the Signal Hypothesis: Early Experiments
In the test of the Signal Hypothesis (which won Blobel the 1999 Nobel Prize in Physiology or Medicine), isolated RER membranes were included with mouse myeloma cell mRNA in cell-free protein synthesis systems. Electrophoretic autoradiographs this time showed that the polypeptides made in vitro in the presence of RER were the same size as the mature, secreted polypeptides. The RER must therefore contain processing activity, i.e., a signal peptidase that removes the signal peptide! The steps of the signal hypothesis that emerged from the experiments of Blobel and his colleagues are illustrated below.
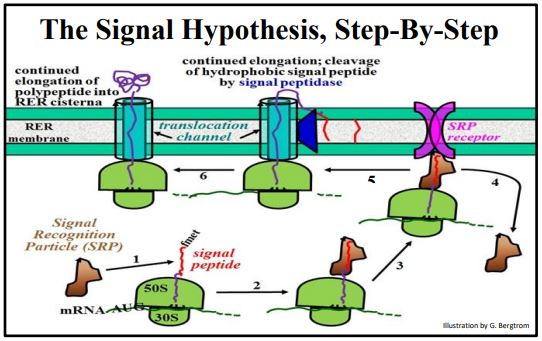
Recall that the synthesis of any protein starts with assembly of a translation initiation complex, followed by polypeptide elongation. During elongation, the growing polypeptide moves through and emerges from a channel, or groove in the large subunit. As the N-terminal signal sequence (i.e., the signal peptide) of a secretory polypeptide emerges from this groove, it interacts with the RER membrane. Beginning at the lower left of the illustration above, the steps of the process are:
- An SRP (signal recognition particle) binds to the hydrophobic signal peptide.
- Elongation stops until the SRP-ribosome complex finds the RER membrane.
- The ribosome-SRP complex binds to an SRP receptor on the RER membrane.
- The SRP detaches from the growing polypeptide chain, to be recycled.
- Translation elongation resumes through a translocation channel; a signal peptidase in the RER membrane catalyzes co-translational hydrolysis of the signal peptide, which remains embedded in the RER membrane.
- Elongation continues and the growing polypeptide begins to fold in the RER.
305 Testing the Signal Hypothesis
306 Details of the Signal Hypothesis
Step 2 above requires that the SRP find and bind to the signal peptide before the nascent polypeptide gets too long and starts to fold into a 3D (tertiary) conformation. It turns out the ribosome itself may keep the signal peptide available by destabilizing electrostatic interactions that would otherwise lead to folding and an undoubtedly incorrect conformation. For more on ribosome involvement in protein folding, check out the link at Protein Folding-Destabilizing One Protein Strand at a Time.
The secretory mechanism just described for eukaryotes has its counterpart in bacteria, which secrete proteins that assist in nutrient scavenging as well as cell wall synthesis. Partially elongated signal peptides guide mRNA-bound ribosomes to the cytoplasmic side of the plasma membrane, where the ribosomes bind and then pass elongating proteins through the plasma membrane into the space between the cell membrane and wall. As the protein exits the cell, a bacterial signal peptidase (SPase) cleaves the signal peptide. Apparently, the mechanism for the secretion of proteins evolved early and since been conserved. As we will see, this mechanism has been further coopted by eukaryotes for packaging proteins into some organelles and into membranes themselves. Some interesting speculations on the evolution of the protein packaging pathway are discussed in the link below.
307 Destinations of Protein Traffic and Evolution of Pathways
Early on, we discovered that antibiotics stop bacterial growth either by disrupting the cell wall or otherwise killing the cells outright. We now know that some antibiotics (e.g., arylomycins) disrupt plasma membrane SPase function, preventing proteins required in the space between the cell wall and membrane from ever making it out of the cell. Once used against Staphylococcus aurease, arylomycins are no longer effective because many strains have become resistant to these antibiotics (click Bacterial Signal Peptidase and Antibiotic Resistance to read about the mechanism of arylomycin resistance). As you may already know, S. aurease is now resistant to many antibiotics, and illness from untreatable infections has its own name, MRSA (Methicillin-Resistant Staph Aurease - dig on your own to see more about methicillin resistance). While named for methicillin resistance, MRSA now describes nearly untreatable S. aurease infections.
B. Synthesis of Membrane-Spanning (Integral) Proteins
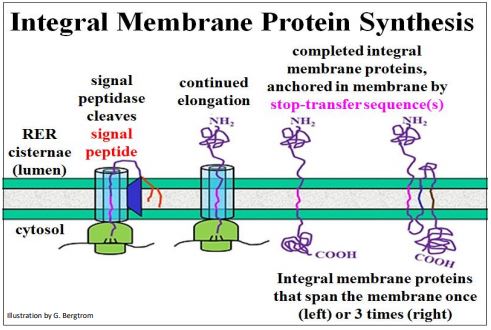
N-terminal signal sequences also guide ribosomes translating integral membrane proteins to the RER. However, before such a protein can pass completely through the membrane, a stop-transfer sequence (a hydrophobic domain within the polypeptide chain) traps the protein in the fatty acid interior of the membrane. Multiple stoptransfer sequences account for transmembrane proteins that span a membrane more than once (below).
C. Moving and Sorting Packaged Proteins to Their Final Destination
Like proteins packaged in RER, those made in the cytoplasm go to different destinations before they become functional. Let’s look at the sorting mechanisms for proteins sequestered by the endomembrane system and those made in the cytoplasm.
1. Traffic on the Endomembrane Highway
We have already seen that, once packaged in the RER cisternae, proteins begin post-translational modification (by e.g., ‘core glycosylation’). Transport vesicles that bud off from the RER carry packaged and membrane proteins to the cis vesicles of the Golgi apparatus. There, vesicle fusion is mediated by the recognition of complementary integral membrane proteins embedded in the two membranes. Later, such packaged proteins are sorted to different organelles or to the plasma membrane. Sorting starts as proteins move from the cis to the trans face of the Golgi vesicles, where specific sorting proteins associate with different packaged proteins in the trans Golgi vesicles. The packaged proteins then sort to vesicles that bud off from trans Golgi stacks. These vesicles move to their final destinations, recognizing and fusing with appropriate membranes. Some events of protein trafficking are animated at Events in Protein Trafficking and summarized in the illustration on the next page.
James E. Rothman, Randy W. Schekman and Thomas C. Südhof won the 2013 Nobel Prize in Physiology or Medicine for their studies of the regulation of vesicle traffic (click 2013 Nobel Prize in Physiology or Medicine for more information). Let’s follow some proteins in and on RER membranes through the cell:
- Transition vesicles carrying their mix of packaged proteins bud off from the RER with the help of COPI and COPII coat proteins, and dissociate from the ribosomes originally attached to them. Transition vesicles however, remain associated with the COP proteins.
- These vesicles fuse with the cis Golgi vesicles, a process also mediated by COP proteins. COPI proteins detach during or after fusion, to be recycled back to the RER .
- Packaged proteins and membrane proteins are further processed as the pass through the Golgi vesicle stack, for example undergoing terminal glycosylation.
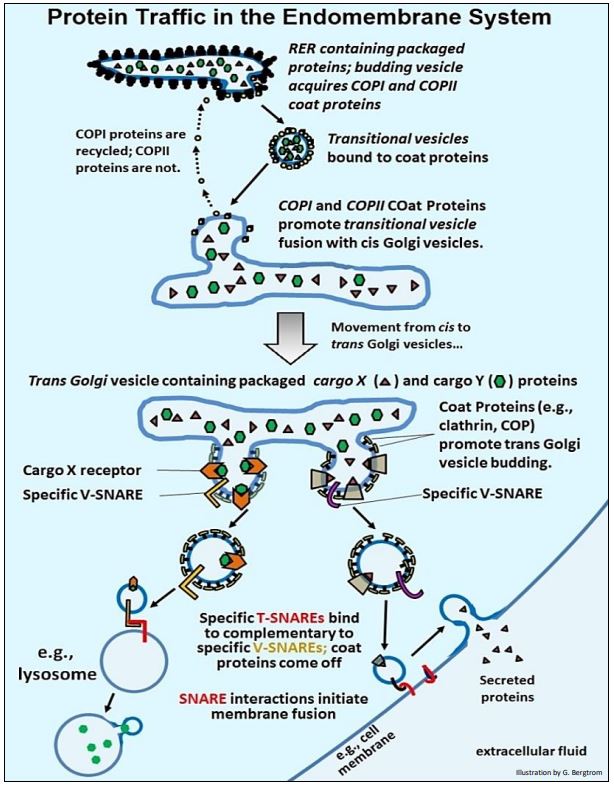
- At the trans face of the Golgi vesicles, cargo receptor proteins in the membranes to bind specific packaged proteins (now called cargo proteins). With the help of clathrin and other COP proteins, cargo protein-bound receptor proteins bud off from the trans Golgi stack. However this time, specific cargo proteins sort to separate vesicles with different cellular or extracellular destinations. These budding vesicles also acquire membrane V-SNARE (for vesicle-SNARE) proteins.
- When V-SNARE proteins on their vesicles bind to complementary T-SNARE (for target-SNARE) proteins on receiving membranes, the membranes fuse.
- Some vesicles follow this pathway, fusing with lysosomes or similar vesicles to stock them with appropriate enzymes and other protein content. Coat proteins come off the fusing vesicle and are recycled, while vesicle contents are transferred into the next vesicle.
- Vesicles containing secretory proteins typically fuse to form larger secretory vesicles. Secretory vesicles can be stored until the cells are signaled to release their contents from the cell. At that point, secretion vesicles fuse with the plasma membrane, releasing their contents to the extracellular fluid. Once again, coat proteins and clathrin dissociate from the secretory vesicle during fusion.
Other players have been left out of this discussion, notably those that hydrolyze nucleotide triphosphates to provide the energy for this protein trafficking. In addition, you might recognize other molecular players such as clathrin that play a role receptor-mediated endocytosis. Maybe that’s not a surprise! After all, endocytosis is, at least partly, molecular traffic in the opposite direction of vesicle formation and secretion.
2. Nuclear Protein Traffic
Almost all proteins are encoded in the nucleus and translated in the cytosol. These include most of those found in nucleus itself, as well as in mitochondria and chloroplasts (see the Endosymbiotic Hypothesis for a description of intraorganelle gene expression). Proteins synthesized in the cytosol destined for these organelles contain oligopeptide traffic signals that direct them to their appropriate destinations. We saw earlier that large molecules (mRNAs, tRNAs) and even whole particles (i.e., ribosomal subunits) cross the nuclear envelope through nuclear pores.
As for proteins headed for the nucleus, nuclear localization signals rich in positively charged amino acids (lysine, proline) enable binding to the negatively charged domain of a nuclear transport receptor protein in the cytosol. This process is illustrated below.
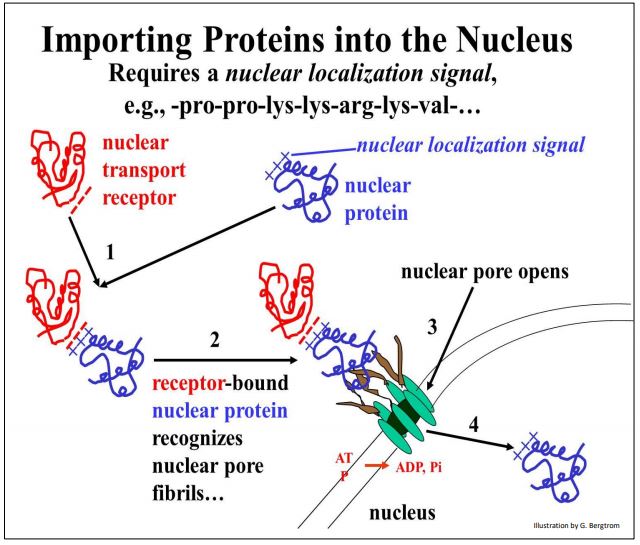
As the complex of the two proteins approach a nuclear pore, it interacts with nuclear pore fibrils, causing the pore to open. The two bound proteins then cross the double membrane of the nuclear envelope where they accumulate against a concentration gradient. This active transport comes from ATP hydrolysis as the nuclear proteins enter the nucleus.
3. Mitochondrial Protein Traffic
Recall that mitochondria contain their own genome and translational machinery. Thus, they transcribe RNAs and translating proteins of their own. However, genes in the nucleus encode many of the proteins found in mitochondria. Import of these proteins into mitochondria is illustrated below.
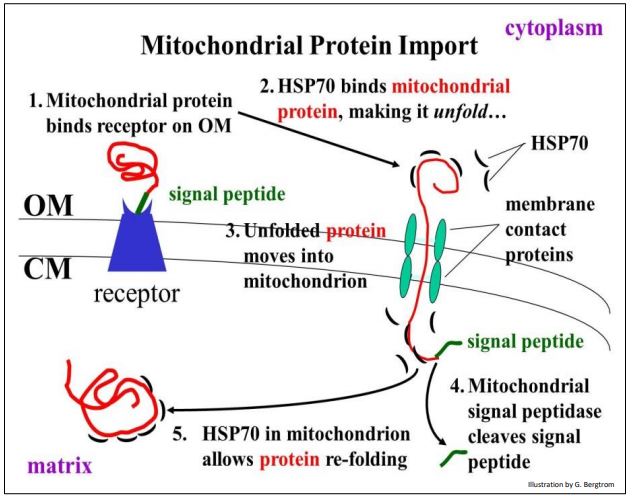
Unlike the co-translational packaging of proteins by the RER, mitochondrial protein transfer is post-translational. This means that mitochondrial proteins formed in the cytoplasm have already folded, assuming a tertiary structure. However, the folded protein exposes an N-terminal signal peptide on its surface that recognizes and binds to a receptor protein at the outer mitochondrial membrane. The receptor protein spans both the mitochondrial outer membrane (OM) and cristal membrane (CM).
The receptor protein delivers the protein to membrane contact proteins that also span both mitochondrial membranes. The membrane contact proteins acts as a channel, or pore, through which the mitochondrial protein will cross into the mitochondrial matrix.
But there is a problem: the folded protein cannot cross the membrane by itself! The entry of a completed mitochondrial protein in the cytoplasm requires a so-called chaperone protein, in this case the HSP70 (heat-shock 70) protein. HSP70 controls unfolding of the mitochondrial protein as it passes into the matrix. Upon removal of the signal peptide by a mitochondrial signal peptidase, another HSP70 molecule resident in the mitochondrion facilitates refolding of the protein into a biologically active shape. Recall that HSPs were initially discovered in heat