
17.2: Membrane Transport
- Page ID
- 16521

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)The first control on the passage of molecules across membranes is the semi-permeable character of the membrane itself. Molecules move in and out of cells in one of three ways: passive diffusion, facilitated transport and active transport.
Only a few small, relatively uncharged molecules can cross a membrane unassisted (i.e., by passive diffusion). Hydrophilic molecules that must enter or leave cells do so with help, i.e., by facilitated transport. Passive and facilitated transport release the free energy inherent in concentration gradients as molecules diffuse across a membrane. In contrast, active transport consumes energy to create concentration gradients of specific solutes. The specificity of facilitated and active transport lies in integral membrane proteins that recognize and bind specific solutes for transport. As you may predict, allosteric regulation of these proteins controls the movement of their target molecules into or out of cells.
Despite its polarity, many believed that the small water molecules crossed membranes without help. Indeed, it does to a limited extent. However, others suspected that given its highly charged polar covalent bonds relative to its small size, water molecules require an assist to get across membranes efficiently. Let’s begin with a closer look at passive diffusion and facilitated diffusion, followed by osmosis (a special case of facilitated diffusion), and finally, at active transport.
A. Passive Diffusion
Passive diffusion is the movement of molecules over time by random motion (also called Brownian motion) from regions of higher concentration to regions of lower concentration. Significant passive diffusion across cellular membranes is limited to a few molecules, mostly gasses like O2, CO2, and N2, that can freely cross the hydrophobic phospholipid barrier. The rapid diffusion of gasses is essential for O2 and CO2 exchange between the alveolar capillaries and cells of the lungs during physiological respiration. O2 and CO2 exchange also occurs in mitochondria during cellular respiration. Diffusion across membranes does not require energy. In fact, diffusion releases energy - recall the movement of protons through the F1 ATPase proton gate that synthesizes ATP during mitochondrial oxidative phosphorylation.
The rate of diffusion of a molecule is dependent only on its own concentration. It is unaffected by the concentration of other molecules. Over time, random motion of solutes within and across compartments results in a dynamic equilibrium for each different solute over time. At equilibrium, solute molecules continue to diffuse across the membrane, but for each molecule moving across in one direction, another molecule of the same solute crosses in the other direction.
B. Facilitated Diffusion of Solutes and Ions
Like passive diffusion, facilitated diffusion is the spontaneous (downhill) passage of molecules or ions across membranes through specific transmembrane proteins. The kinetics of passive and facilitated diffusion reveals the differences between the two processes. To understand the latter, recall that the rate of enzyme catalysis is saturable. That is, as the concentration of substrate is increased, the rate of the catalyzed reaction approaches a maximum (Vmax), when all enzyme molecules in solution are bound to substrate molecules. The same saturation phenomenon applies to facilitated transport – the rate of solute movement across a membrane is directly proportional to the number of transport proteins in the membrane.
The kinetics of passive and facilitated diffusion are illustrated by the graph shown below
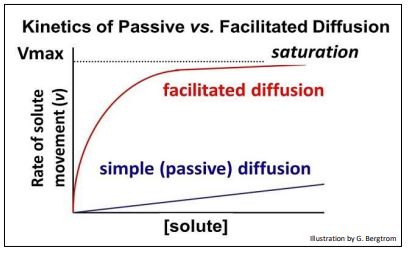
Perhaps you see another similarity between facilitated diffusion and enzyme catalysis in this graph! Relative rates of facilitated diffusion are typically rapid, compared to those of passive diffusion. This is because the allosteric changes that accompany facilitated transport are rapid, just as they are during enzyme catalysis. There are three kinds of facilitated transport of solutes (below).
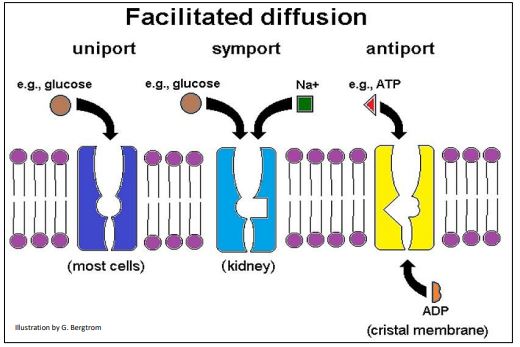
The GLUT protein (glucose transporter) protein shown above (left) allows glucose uniport, the specific transport of a single substance in or out of cells. Kidney cells have glucose transporters that symport (couple) the simultaneous movement of glucose and sodium ions; SGLT (Sodium-GLucose Transporter) serves a similar function in small intestine cells, enabling absorption of dietary glucose and sodium. Antiport (above, right) allows the specific exchange of molecules across a membrane. In the example shown, ATP leaves the mitochondrial matrix, crossing the cristal membrane at the same time as ADP enters the matrix.
Whether by uniport, symport or antiport, each solute will independently cross a membrane down its concentration gradient, moving from higher concentration to where it is at a lower concentration. Recall that diffusion along a gradient releases free energy that depends on relative concentrations of the solutes.
Proteins mediating facilitated diffusion are of two kinds: carrier proteins and channel proteins. Carrier proteins allow solute transport. Ions, with their high charge-to-mass ratio, need help to cross the hydrophobic membrane barrier; this is the job of channel proteins that essentially serve as ion pores.
Like all transporter proteins, both carrier and channel proteins undergo allosteric change during transport. They are also typically subject to allosteric regulation, rather than being in a constant ‘open’ state. Examples of facilitated diffusion are considered in more detail below.
1. Carrier Proteins
When a carrier protein binds a solute that must cross the membrane, it undergoes an allosteric change (illustrated below). During transport, the carrier protein undergoes another change in shape. When the solute reaches the other side of the membrane, it no longer has a high affinity for the carrier protein. After release of the solute, a final allosteric change restores the original conformation of the transport protein. These sequential conformational changes are illustrated on the next page.
Any given carrier protein is specific for a single solute, or at most a single family of closely related solutes. We just saw the GLUT1 transporter carrier protein that allows glucose (but not fructose or ribose!) to cross cell membranes. Different specific carrier proteins facilitate the transport of amino acids and other charged solutes across cell membranes. Once again, molecules that indicate cell status (i.e., a need to import or export solute) are allosteric effectors that regulate carrier proteins. The regulation of glucose transport into cells by insulin is a perfect
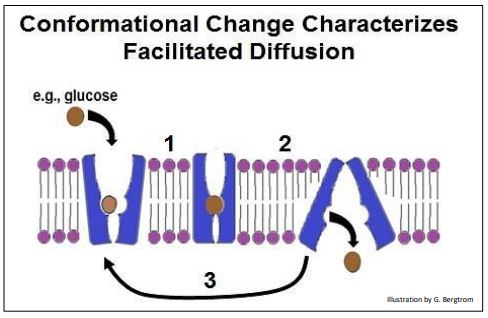
example. One consequence of insulin released during a meal (or just in anticipation of a meal) is the stimulation of glucose transporters to take up glucose. An inability of those transporters to respond to insulin accounts in part for Type II (adult onset) diabetes.
Water gets across membranes by osmosis (we’ll look more closely at how osmosis affects cells in a moment). We noted that small amounts of water could cross the phospholipid bilayer unassisted. Water can also cross a membrane incidentally, when ions flow through their channel proteins. But most osmosis involves facilitated diffusion mediated by aquaporins. Some aquaporins only transport water. Others have evolved to co-facilitate the transport of glucose (see above), glycerol, urea, ammonia, carbon dioxide and even ions (protons) along with water. Like other carrier proteins, aquaporins are allosterically regulated to allow cells to meet their specific water balance requirements. So fundamental was the understanding of water balance that the discovery of aquaporins earned Peter Agre a Nobel Prize in Chemistry in 2003. Since Agre’s discovery (in 1992), several genetic diseases have been linked to aquaporin gene mutations.
Kidney cells are critically involved in vertebrate water balance and have many aquaporins in their membranes. In a rare form of diabetes, abnormal aquaporins cause the kidneys to excrete unusually large volumes of water. In another example, aquaporin gene mutations lead to the development of cataracts in both eyes. Since their initial discovery, aquaporins have been described in bacteria and plants. To learn more, click Aquaporins.
2. Ion Channels
Allosteric regulation of ion channel proteins controls ion homeostasis in blood and extracellular fluids within narrow limits. Often, multiple integral proteins contribute to the formation of an ion channel. When stimulated, channel proteins rearrange to open a pore allowing specific ion transport. Some ion channels, like the glucosesodium ion symport system noted above, mobilize the energy of diffusion of one solute (an ion in this case) to rapidly transport another solute through the same channel (acting like an ion channel and a carrier protein). Finally, ion channels are responsible for the excitability of cells, where Na+ , K+ and Ca++ channels collaborate in ion movements into and out of cells leading to neuronal or muscle cell responses (more shortly!)
C. Osmosis
Osmosis, the diffusion of water across membranes from lower to higher solute concentrations, is an essential activity. It allows cells to use water to maintain cellular integrity or to adapt to changes in the solute composition in the extracellular environment. Osmosis across cellular membranes relies on the facilitated transport of water by aquaporins. The passive diffusion of water molecules, can be demonstrated with an artificial (e.g., dialysis) membrane. Water will cross such a membrane if solute concentrations are higher on one side of the membrane. Water crosses the membrane “trying” to equalize the solute concentrations on both sides of the membrane. In effect, water movement is from the side of a membrane where the free water molecule concentration is higher (i.e., where the concentration of solute is lower) to the side where the concentration of free water is lower (i.e., where the concentration of solute is higher).
1. Osmosis in Plant and Animal Cells
We could present this section in the context of free water concentrations, but we will do so in the more familiar terms of solute concentrations. Osmosis affects plant and animal cells according to the same principles, but with different effects. First, let us consider the effect of different experimental solute concentrations on animal cells, illustrated on the next page.
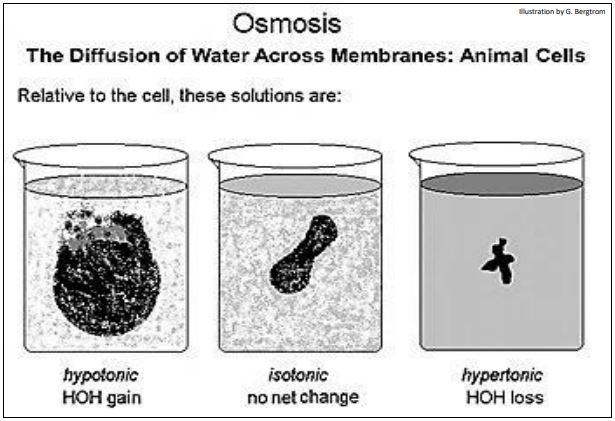
If the solute concentration inside and outside the cell is the same, there is no net movement into or out of the cells. The extracellular medium and cytosol are said to be isotonic to each other. When water diffuses into the cells from a low solute medium, the medium is said to by hypotonic to (less concentrated than) the cytosol. In this case, movement of water into a cell lowers the cytosol solute concentration. Animal cells swell and burst in a hypotonic solution. In hypertonic solutions (with a higher solute concentrations than the cytosol), animal cells shrivel up as water leaves the cell. From this brief description, you should conclude that water crosses from the hypotonic to the hypertonic side of a membrane.
As with animal cells, exposure of plant cells to hypotonic or hypertonic solutions causes the same directional water movements, but with some key differences due to their cell walls. In hypotonic solutions, water enters plant cells, moving into the cytosol and then into water vacuoles called tonoplasts. This results in higher osmotic pressure (water pressure) in the tonoplasts. The expanding tonoplast creates turgor pressure, compressing the cytosol against the cell wall. Rather than bursting, the cells and plant tissues stiffen and become turgid. Since water cannot enter plant cells indefinitely, water stops entering the cells when the osmotic pressure outside the cells and the turgor pressure inside the cells are at equilibrium. You encountered this phenomenon if you have ever over-watered houseplants. The stiffened leaves and stems become brittle and are easily snapped or broken. In hypertonic medium, plant cells (like animal cells) lose water, resulting in plasmolysis. This is the effect of shrinkage of the plasma membrane. However, the plasma membrane remains tightly attached to the plant cell wall at several points. You may have seen under-watered plants with floppy or droopy stems and leaves. These have become flaccid due to loss of water and thus the loss of turgor pressure needed to keep leaves and stems upright. The effects of different solutions on plant cells are illustrated below
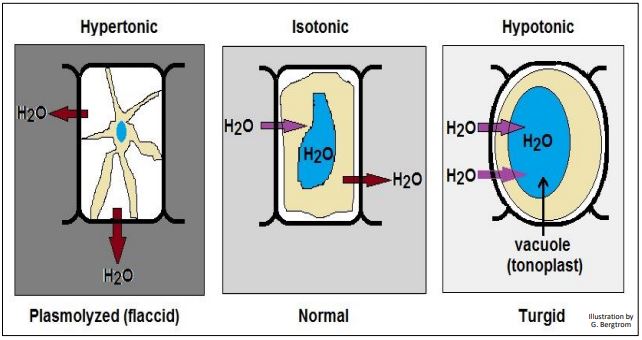
Formally, osmotic or turgor pressure is defined as the force per unit area (i.e., pressure) required to prevent the passage of water across a semipermeable membrane from a hypotonic to a hypertonic solution
2. Osmosis in Plant Life
While individual plant cells respond to changes in solute concentrations, these changes are rapidly communicated to adjacent cells through plasmodesmata. These structures connect the plasma membranes of adjacent cells through their cell walls, allowing rapid, direct sharing of physical and chemical information.
A plasmodesma is illustrated below.
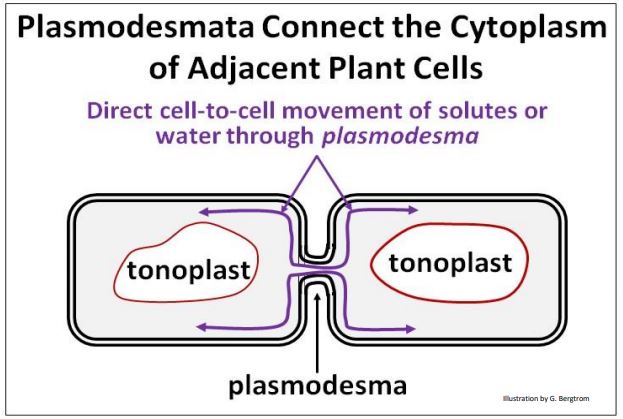
In this way, effects on osmotic pressure in a few cells created by changes in water availability are transmitted to adjacent cells, affecting turgor pressure in those cells and, ultimately, in plant tissues.
Finally, plant life depends on water! Recall that plant cells require a continual supply of water for use in photosynthesis, to provide hydrogen to reduce CO2 to glucose. Photosynthesis as well as the loss of excess water from plant tissues (especially leaves) by transpiration lowers cellular osmotic pressure. As water moves up from the roots to replace water used and lost by leaf cells, the osmotic pressure drops in the fine root hair cells (with their high surface area). This draws water into the cells and roots by osmosis. Thus, osmotic pressure is the main force driving water into plants and, defying gravity, moving it up from the roots to the rest of the plant
3. Osmosis in Animal Life
Changes in osmotic environment can stress or kill an organism. For example, freshwater organisms (protozoa or fish) placed in sea water will die. Likewise, saltwater fish placed in freshwater. But organisms can osmoregulate (control the osmotic pressure in their cells), at least to a point. Paramecium for example, expels fresh water to prevent bursting as it takes on water. This is accomplished by a contractile vacuole (shown below).

Water constantly enters these freshwater protists because the solute concentration in the cytosol is always higher than the freshwater water they live in. To cope with a constant uptake of water, their contractile vacuoles collect excess water and then contract to expel the water. At a high-energy cost, Paramecia constantly pump water out of the cell to maintain water balance (i.e., correct osmotic pressure). Another protist strategy for coping with change in environmental solute concentrations (salinity) is to pump salts (or suitable salt solute substitutes in or out of the cell, as needed (For some details, see Protist Osmoregulation Genes Acquired by Eukaryotes from Bacteria by Horizontal Gene Transfer).
Larger organisms like freshwater fish cope with their hypotonic environment by urinating a lot! At the other end of the spectrum, salt-water fish cope with the high solute concentration of solutes (salts) in their environment by excreting excess salt. Salmon spend time in seawater growing to maturity and later swim upstream in fresh water to spawn. You can imagine how salmon and similar organisms have to osmoregulate to adapt to their changing, very different environments. In this case, osmoregulation begins when hormonal changes respond to changes in living circumstance and dictate a compensatory response.
4. Summing Up
Osmosis is the movement of water across membranes to where solutes are at high concentration. At the same time, solutes that can diffuse across membranes move in or out of cells towards where they are at lower concentration, either passively, or by facilitated diffusion. We have evolved different facilitated transport proteins specific for different proteins. Finally, most water crosses membranes by facilitated diffusion through aquaporin proteins that serve as pores in cellular membranes.
D. Active Transport
Excitability (adaptation) is another of the defining properties of life. This property of all cells is based on chemical and electrical reactivity. Neurotransmitters released at a synapse cross the synaptic cleft from a “sending” neuron to a responding cell (another neuron or a muscle cell). The neurotransmitter binds to receptors on the responding cell resulting in a membrane depolarization, a rapid change in the electrical potential difference across the cell membrane. While responses to neurotransmitters occur in fractions of a second, all cells are responsive, albeit not always as fast as neurons or muscle cells. Changes in membrane polarity of any cell depend on unequal concentrations of ions inside and outside cells. Cells at rest typically have a higher [K+] in the cytosol and higher [Cl-] and [Na+] outside the cell (below).
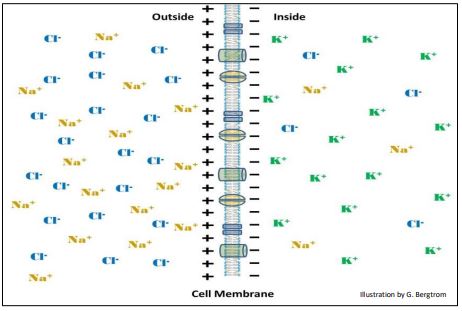
These ionic differences across membranes are what enable such cells as neurons and muscle to respond to chemical and other (e.g., electrical) signals. Thus, cells have a resting potential, shown here with plus and minus signs on opposite sides of the membrane. The measured resting potential (difference in charge or potential difference) of most cells is typically between -50mv to -70mv. Disturbance of the resting potential (i.e., membrane depolarization), results from a flow of ions across membranes. Resting potentials sustained by ion gradients permit physiological response to chemical or other signals.
Resting potentials change when cells are excited, as well as by normal, but non-functional ion leakage. Whether incidental or intentional, the correct ion balance must be restored and maintained. This is accomplished by the active transport of ions across the membrane. This energetically unfavorable process requires an input of free energy, typically from ATP hydrolysis. The Na+/K+ pump is an active transport protein complex linked to ATPase activity. Next, we consider ion flow during cell excitation and how ion pumps work.
Let’s begin by looking at the allosteric changes that occur when the Na+ /K+ pump works to restore and maintain ion gradients (illustrated on the next page). In operation, the ATPase domain of the Na+ /K+ pump hydrolyzes ATP, leaving a phosphate attached to the pump and inducing the first of several allosteric changes in the pump proteins (No. 1, above). In its new conformation, the pump binds three Na+ ions, causing a second conformational change that in turn releases the Na+ ions into the extracellular fluid (No. 2). The release of Na+ ions outside the cell causes a third allosteric change (No. 3), after which two K+ ions from the extracellular fluid are able to bind to the pump protein. K+ binding causes the hydrolysis of the phosphate from the pump protein, returning the pump proteins to their original conformation (No. 4) and releasing the two K+ ions into the cytosol. The Na+ /K+ pump is ready for action again!
295 Potassium Leakage Helps to Maintain Cellular Resting Potentials
296 Active Transport by the Sodium/Potassium Pump
For his discovery of the ATPase-powered sodium/potassium pump and and his studies of how it works to maintain intracellular ion balance, Jens C. Skou earned a share of the Nobel Prize in Chemistry 1997. You can read more about Jens C. Skou at https://www.nobelprize.org/prizes/chemistry/1997/skou/auto-biography/
