
3.1.2: Plant Cell Structure
- Page ID
- 27731

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)- Describe the structures that all cells share in common.
- State the role of the plasma membrane.
- Describe the structures of eukaryotic cells.
- Summarize the functions of the major cell organelles.
Components of All Cells
All cells contain these same four components: 1. plasma (cell) membrane, a phospholipid bilayer with a mosaic of proteins, which functions as a barrier between the cell and its environment. 2. cytoplasm, the region between the region of DNA and plasma membrane, and the cytosol, a fluid, jelly-like region inside the cell where chemical reactions take place. 3. DNA, the heredity information of cells, which can be found in a nucleus of eukaryotic cells and the a nucleoid region of prokaryotic cell. 4. ribosomes, or protein-synthesizing structures composed of ribosomes and proteins. These structures can be found on the image of the plant cell (Figure \(\PageIndex{1}\)).

The Plasma Membrane
Both prokaryotic and eukaryotic cells have a plasma membrane (Figure \(\PageIndex{2}\)), a phospholipid bilayer with embedded proteins that separates the internal contents of the cell from its surrounding environment. A phospholipid is a lipid molecule with two fatty acid chains and a phosphate-containing group. The plasma membrane controls the passage of organic molecules, ions, water, and oxygen into and out of the cell. Wastes (such as carbon dioxide and ammonia) also leave the cell by passing through the plasma membrane. The plasma membrane are semi-permeable and allow small and/or non-polar molecules to pass through. Water, being small, can pass through the membrane and will move from an area of low solute concentration to an area of high solute concentration by the process of osmosis.
(Figure \(\PageIndex{2}\)): The eukaryotic plasma membrane is a phospholipid bilayer with proteins and cholesterol embedded in it.
The Cytoplasm
The cytoplasm is the cell's entire region between the plasma membrane and the nuclear envelope (a structure we will discuss shortly). It is comprised of organelles suspended in the gel-like cytosol, the cytoskeleton, and various chemicals (Figure \(\PageIndex{1}\)). Even though the cytoplasm consists of 70 to 80 percent water, it has a semi-solid consistency, which comes from the proteins within it. However, proteins are not the only organic molecules in the cytoplasm. Glucose and other simple sugars, polysaccharides, amino acids, nucleic acids, fatty acids, and derivatives of glycerol are also there. Ions of sodium, potassium, calcium, and many other elements also dissolve in the cytoplasm. Many metabolic reactions, including protein synthesis, take place in the cytoplasm.
DNA
In eukaryotic cells, the DNA is typically housed in a nucleus (plural = nuclei), the most prominent organelle in a cell (Figure \(\PageIndex{1}\). This organelle directs the synthesis of ribosomes and proteins. Let’s look at it in more detail (Figure \(\PageIndex{3}\)).
(Figure \(\PageIndex{3}\)): The nucleus stores chromatin (DNA plus proteins) in a gel-like substance called the nucleoplasm. The nucleolus is a condensed chromatin region where ribosome synthesis occurs. We call the nucleus' boundary the nuclear envelope. It consists of two phospholipid bilayers: an outer and an inner membrane. The nuclear membrane is continuous with the endoplasmic reticulum. Nuclear pores allow substances to enter and exit the nucleus.
The Nuclear Envelope
The nuclear envelope is a double-membrane structure that constitutes the nucleus' outermost portion (Figure \(\PageIndex{3}\). Both the nuclear envelope's inner and outer membranes are phospholipid bilayers.
The nuclear envelope is punctuated with pores that control the passage of ions, molecules, and RNA between the nucleoplasm and cytoplasm. The nucleoplasm is the semi-solid fluid inside the nucleus, where we find the chromatin and the nucleolus.
Chromatin and Chromosomes
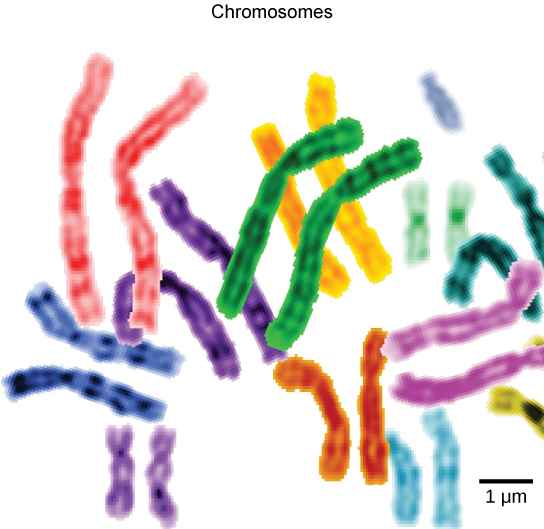
To understand chromatin, it is helpful to first explore chromosomes, structures within the nucleus that are made up of DNA, the hereditary material. You may remember that in prokaryotes, DNA is organized into a single circular chromosome. In eukaryotes, chromosomes are linear structures. Every eukaryotic species has a specific number of chromosomes in the nucleus of each cell. For example, in humans, the chromosome number is 46, while in fruit flies, it is eight. Chromosomes are only visible and distinguishable from one another when the cell is getting ready to divide. When the cell is in the growth and maintenance phases of its life cycle, proteins attach to chromosomes, and they resemble an unwound, jumbled bunch of threads. We call these unwound protein-chromosome complexes chromatin (Figure \(\PageIndex{4}\). Chromatin describes the material that makes up the chromosomes both when condensed and decondensed.

The Nucleolus
We already know that the nucleus directs the synthesis of ribosomes, but how does it do this? Some chromosomes have sections of DNA that encode ribosomal RNA. A darkly staining area within the nucleus called the nucleolus (plural = nucleoli) aggregates the ribosomal RNA with associated proteins to assemble the ribosomal subunits that are then transported out through the pores in the nuclear envelope to the cytoplasm.
Ribosomes
Ribosomes are the cellular structures responsible for protein synthesis. They are not organelles. They can be small dot-like structures that float freely in the cytoplasm (known as free ribosomes) or they may be attached to the plasma membrane's cytoplasmic side or the endoplasmic reticulum's cytoplasmic side and the nuclear envelope's outer membrane, and called attached ribosomes (Figure \(\PageIndex{1}\)). Ribosomes are large protein and RNA complexes consisting of two subunits, a large and a small (Figure \(\PageIndex{5}\). Ribosomes receive their “orders” for protein synthesis from the nucleus where the DNA transcribes into messenger RNA (mRNA). The mRNA travels to the ribosomes, which translate the code provided by the sequence of the nitrogenous bases in the mRNA into a specific order of amino acids in a protein. Amino acids are the building blocks of proteins.
Figure \(\PageIndex{5}\): A large subunit (top) and a small subunit (bottom) comprise ribosomes. Note that the large subunit sits on top of the small subunit. During protein synthesis, ribosomes assemble amino acids into proteins using information from a chain of mRNA. The tRNA carries amino acids to the ribosome, where the arrangement is made.
Because protein synthesis is an essential function of all cells (including enzymes, hormones, antibodies, pigments, structural components, and surface receptors), there are ribosomes in practically every cell. Ribosomes are particularly abundant in cells that synthesize large amounts of protein. For example, the pancreas is responsible for creating several digestive enzymes and the cells that produce these enzymes contain many ribosomes. Thus, we see another example of form following function.
Components Unique to Eukaryotic Cells
All cells contain DNA, as described above. However, plant cells, which are eukaryotic, contain organelles and a nucleus while prokaryotic cells do not possess organelles or a membrane bound nucleus. We will start by going over the structures that are unique to all eukaryotic. Next, we will go over structures unique to plant cells.
Endomembrane System
The endomembrane system (endo = “within”) is a group of membranes and organelles (Figure \(\PageIndex{6}\)) in eukaryotic cells that works together to modify, package, and transport lipids and proteins. It includes the nuclear envelope, lysosomes, and vesicles, the tonoplast (see below), and the endoplasmic reticulum and Golgi apparatus. Although not technically within the cell, the plasma membrane is included in the endomembrane system because, as you will see, it interacts with the other endomembranous organelles. The endomembrane system does not include either mitochondria or chloroplast membranes.
Visual Connection
If a peripheral membrane protein were synthesized in the lumen (inside) of the ER, would it end up on the inside or outside of the plasma membrane?
The Endoplasmic Reticulum
The endoplasmic reticulum (ER) (Figure \(\PageIndex{6}\)) is a series of interconnected membranous sacs and tubules that collectively modifies proteins and synthesizes lipids. They are formed as an extension of the nuclear membrane and fold out towards the cytoplasm. The two functions of the ER take place in separate areas: the rough ER and the smooth ER, respectively.
The rough endoplasmic reticulum (RER) can be found has ribosomes along its surface, and the proteins they create are either secreted or incorporated into membranes in the cell. The smooth endoplasmic reticulum (SER) is continuous with the RER but has few or no ribosomes on its cytoplasmic surface (Figure \(\PageIndex{6}\)). SER functions include synthesis of carbohydrates, lipids, and steroid hormones; detoxification of medications and poisons; and storing calcium ions.
Vesicles
Transport vesicles, composed of endomembrane system material, bud off the from the RER, carrying material into the Golgi Apparatus, the next component of the endomembrane system.
Golgi Apparatus
The lipids or proteins within the transport vesicles still need sorting, packaging, and tagging so that they end up in the right place. Sorting, tagging, packaging, and distributing lipids and proteins takes place in the Golgi apparatus (also called the Golgi body), a series of flattened membranes (Figure \(\PageIndex{6}\)).
We call the side of the Golgi apparatus that is closer to the ER the cis face. The opposite side. closer to the plasma membrane, is the trans face. The transport vesicles that formed from the ER travel to the cis face of the Golgi, fuse with it, and empty their contents into the Golgi apparatus' lumen. As the proteins and lipids travel through the Golgi, they undergo further modifications that allow them to be sorted. The most frequent modification is adding short sugar molecule chains. These newly modified proteins and lipids then tag with phosphate groups or other small molecules in order to travel to their proper destinations.
Finally, the modified and tagged proteins are packaged into secretory vesicles that bud from the Golgi's trans face. While some of these vesicles deposit their contents into other cell parts where they will be used, other secretory vesicles fuse with the plasma membrane and release their contents outside the cell.
In plant cells, the Golgi apparatus has the additional role of synthesizing polysaccharides, some of which are incorporated into the cell wall and some of which other cell parts use.
Cytoskeleton
The cellular skeleton is a collection of protein filaments within the cytoplasm. Microtubules are key organelles in cell division, they form the basis for cilia and flagella. Plant cells do not have cilia, which are short projections from the cell that function in movement, but the sperm cells of early diverging plants, like bryophytes and seedless vascular plants, have flagella. These are long projections that function in movement. Microtubules are also are guides for the construction of the cell wall, and cellulose fibers are parallel due to the microtubules. The movement in microtubules is based on tubulin-kinesin interactions. In contrast, the movement of microfilaments is based on actin-myosin interactions. Microfilaments guide the movement of organelles within the cell.
Mitochondria
Mitochondria (singular = mitochondrion) are often called the “powerhouses” or “energy factories” of a cell because they are responsible for making a nucleic acid called adenosine triphosphate (ATP), the cell’s main energy-carrying molecule. ATP represents the short-term stored energy of the cell. Cellular respiration is the process of making ATP using the chemical energy found in glucose and other nutrients. In mitochondria, this process uses oxygen and produces carbon dioxide as a waste product. In fact, the carbon dioxide that you exhale with every breath comes from the cellular reactions that produce carbon dioxide as a byproduct.
In keeping with our theme of form following function, it is important to point out that muscle cells have a very high concentration of mitochondria that produce ATP. Your muscle cells need a lot of energy to keep your body moving. When your cells don’t get enough oxygen, they do not make a lot of ATP. Instead, the small amount of ATP they make in the absence of oxygen is accompanied by the production of lactic acid.
Mitochondria are oval-shaped, double membrane organelles (Figure \(\PageIndex{7}\)) that have their own ribosomes and DNA. Each membrane is a phospholipid bilayer embedded with proteins. The inner layer has folds called cristae. The area surrounded by the folds is called the mitochondrial matrix. The cristae and the matrix have different roles in cellular respiration.

Peroxisomes
Eukaryotic cells frequently have smaller vesicles including peroxisomes which, among other functions, help in photosynthesis in plant cells. In addition, many plant cells accumulate lipids as oil drops located directly in cytoplasm. Peroxisomes are small, round organelles enclosed by single membranes. They carry out oxidation reactions that break down fatty acids and amino acids. They also detoxify many poisons that may enter the body. (Many of these oxidation reactions release hydrogen peroxide, H2O2, which would be damaging to cells; however, when these reactions are confined to peroxisomes, enzymes safely break down the H2O2 into oxygen and water.) For example, alcohol is detoxified by peroxisomes in liver cells. Glyoxysomes, which are specialized peroxisomes in plants, are responsible for converting stored fats into sugars.
Components Unique to Plant Cells
The following structures are found exclusively in plant cells and are absent in animal cells.
Cell wall
Though a cell wall is commonly found in prokaryotes and fungi as well as plants, their diversity is due to convergent evolution, not common ancestry, when it comes to these three groups of organisms. Plant cell walls are composed of cellulose are an excretion found outside the plasma membrane. They serve as a covering that provides structural support and gives shape to the cell.
Central Vacuole
The central vacuole is a large, membrane-bound structure that fills much of the plant cell. The membrane surrounding the central vacuole is called the tonoplast. The central vacuole plays a key role in regulating the cell’s concentration of water in changing environmental conditions. Have you ever noticed that if you forget to water a plant for a few days, it wilts? That’s because as the water concentration in the soil becomes lower than the water concentration in the plant, water moves out of the central vacuoles and cytoplasm (Figure \(\PageIndex{8}\)). As the central vacuole shrinks, it leaves the cell wall unsupported. This loss of support to the plant's cell walls results in the wilted appearance. The central vacuole also supports the cell's expansion. When the central vacuole holds more water, the cell becomes larger without having to invest considerable energy in synthesizing new cytoplasm. Lastly, central vacuoles store nutrients, accumulate ions, or become a place to store wastes.

Plastids
Plastids are a group of storage organelle found in plants and algae. Chloroplasts are a type of plastid that store chlorophyll and other pigments for photosynthesis. Chromoplasts are plastids that store orange or yellow pigments, found in plants and fruit such as bell peppers. They are rich in carotenes and xanthophyls. Amyloplasts store starch and can be found in plants such as potato tubers, carrot roots, sweet potato roots, and grass seeds.
Chloroplasts store their pigments in interconnected sacs called thylakoids (Figure \(\PageIndex{9}\)). These sacs are often found in stacks called grana (singular granum). The fluid portion of the double membraned chloroplast is called the stroma. Because the thylakoid stores chlorophyll a, b, and accessory pigments, it is the main region for the first reaction of photosynthesis, where sunlight is used to create molecular energy. In the stroma, the products of the first reaction are used to produce organic molecules such as glucose. The combination of these reactions allow these autotrophic organisms to produce their own organic food.
The chloroplast, like the mitochondria, contains its own DNA, ribosomes, and is double membraned.

Evolution Connection- Endosymbiosis
We have mentioned that both mitochondria and chloroplasts contain DNA and ribosomes. Have you wondered why an organelle would have its own DNA and ribomosome?
The Endosymbiosis Theory explains:
Symbiosis is a relationship in which organisms from two separate species depend on each other for their survival. Endosymbiosis (endo- = “within”) is a mutually beneficial relationship in which one organism lives inside the other. Endosymbiotic relationships abound in nature. For example, there are microbes that produce vitamin K living inside the human gut. This relationship is beneficial for us because we are unable to synthesize vitamin K. It is also beneficial for the microbes because they are protected from other organisms and from drying out, and they receive abundant food from the environment of the large intestine.
Scientists have long noticed that bacteria, mitochondria, and chloroplasts are similar in size. We also know that bacteria have DNA and ribosomes, just as mitochondria and chloroplasts do. Scientists believe that host cells and bacteria formed an endosymbiotic relationship when the host cells ingested both aerobic and autotrophic bacteria (cyanobacteria) but did not destroy them. Through many millions of years of evolution, these ingested bacteria became more specialized in their functions, with the aerobic bacteria becoming mitochondria and the autotrophic bacteria becoming chloroplasts.
Attributions
Curated and authored by Kammy Algiers using the following sources:
- 4.1 Studying Cells and 4.3 Eukaryotic cells from Biology 2e by OpenStax (licensed CC-BY). Access for free at openstax.org.
- 2.2 Mitochondria and Chloroplasts, 2.3 Cell wall, Vacuoles, and Plasmodesmata, and 2.4 Other Parts of the Cellfrom Introduction to Botany by Alexey Shipunov (public domain)