
2.6.3.1: Flowers
- Page ID
- 37015

Learning Objectives
- Identify the components of a flower and to which whorl each belongs.
- Write and interpret floral formulas.
- Differentiate between flowers and inflorescences.
- Explain the difference between raceme-based and cymose inflorescences.
Flowers are sets of highly modified leaves that function to attract a pollinator or, if no animal pollinator is used, to optimize spore dispersal in some way. Over the course of evolutionary history and coevolution, this has lead to an incredible diversity of shape, size, color, smell, and just about any other characteristic you can think of. Because most plants are angiosperms and because flowers are often so diverse, learning the terminology to describe flowers is a major step in learning to identify plants.
The modified leaves in flowers are called sepals, petals, stamens, and carpels (Figure \(\PageIndex{1}\)). These components are arranged in whorls and attach to an area called the receptacle, which is at the end of the stem that leads to the flower. This stem is called the peduncle. In the case of an inflorescence, where multiple florets are produced in place of a single flower, the stems leading to the florets are called pedicels (Figure \(\PageIndex{2}\)).



The general characters that a flower has are whorl morphology, sex, merosity, symmetry, and the position of the gynoecium. Merosity is simply the number of parts in each whorl of a plant structure, whether it is the number of sepals, petals in a corolla, or the number of stamens. Each of these characters will be discussed below.
Whorls
The outermost whorl of a flower is called the calyx and is composed of sepals. Inside the calyx is the corolla, which is composed of petals. The sepals are often smaller and less colorful than the petals, but this general rule can be misleading. For example, lilies and tulips have identical sepals and petals (called tepals, these can be seen in the florets in Figure \(\PageIndex{2}\)). The only way you can distinguish between them is by location: Which whorl is on the outside?
The Perianth: Calyx and Corolla
Together, the calyx and corolla are called the perianth (peri- meaning around, anth- meaning flower; Figure \(\PageIndex{3}\)). The calyx is the outermost whorl of the flower. In most coses, the sepals are not showy--lacking bright colors and typically smaller--and instead serve a protective function in the developing flower. The corolla is the whorl just within the calyx. Petals tend to be the showy part of the flower, specialized for attracting animal pollinators.

Reproductive Whorls: Androecium and Gynoecium
Inside the perianth is the androecium (house of man), a whorl composed of stamens. Each stamen has a long filament holding up pollen sacs called anthers (Figure \(\PageIndex{4-5}\)). Each lobed anther contains microsporangia, within which meiosis of the diploid microspore mother cells in the anther produces four haploid microspores. Each of these develops into a pollen grain consisting of two cells: a larger vegetative cell (the tube cell), inside of which is a a smaller germ cell (also called the generative cell). At some point, depending on the species, the germ cell divides by mitosis to produce 2 spermatia.


Inside the androecium is the gynoecium (house of woman), which is composed of carpels. Each carpel has an ovary at the base where ovules are housed. A style emerges from the ovary and is topped by the stigma (Figure \(\PageIndex{6}\)), where pollen is deposited. A carpel consists of a single ovary, style and stigma. Often several carpels are fused into a single structure, referred to as a pistil.
Within the ovule, a megasporangium produces a megaspore mother cell. Meiosis of the megaspore mother cell in each ovule produces 4 haploid cells, a large megaspore and 3 smaller cells that disintegrate. This megaspore develops into the megagametophyte, all within the ovule. Pollen grains land on the stigma and must grow a tube down the style to reach the ovule and complete fertilization.

Incomplete Flowers
Most flowers are composed of four whorls. If all whorls are present, a flower is said to be both complete and perfect. If any whorl is missing, the flower is incomplete (Figure \(\PageIndex{7}\)). If one of those missing whorls is either the androecium (pollen-producing) or gynoecium (seed-producing), the flower is also imperfect (Figure \(\PageIndex{8}\)).

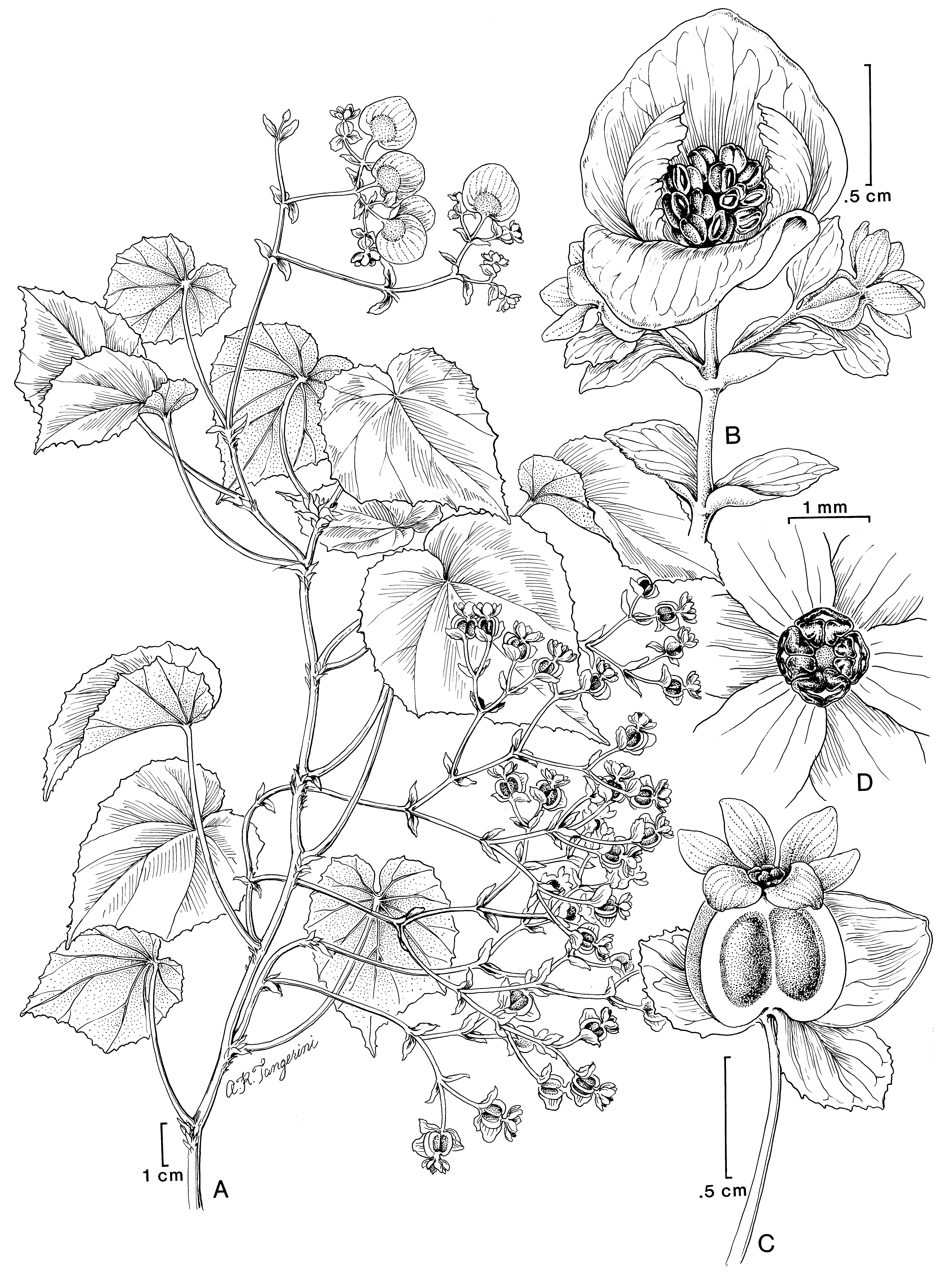
Evolution seems to favor (and be favored by) genetic variability. Genetic variability is promoted by outbreeding - sexual reproduction between genetically dissimilar parents. Just why sexual reproduction is so popular throughout the world of living things is still a hotly-debated question, but the fact remains.
Plants, being anchored in position, have a special problem in this regard. Many employ the services of animals (e.g., insects, birds, bats) to transfer pollen from plant to plant. But if the flowers have both sex organs, what is to prevent the pollen from fertilizing its own eggs? Plants have evolved a variety of solutions. One of these is to produce imperfect flowers.
There are two types of imperfect flowers: staminate flowers contain only an androecium (Figure \(\PageIndex{9}\)), and carpellate (or pistillate) flowers have only a gynoecium. Monoecious plants have both types of imperfect flower on the same plant. Dioecious plants have imperfect flowers on separate plants; that is, some plants are male, some female. Examples include willows, poplars, and the date palm. Most dioecious plants use an X-Y system of of sex determination like that in mammals. However, a few species use an X-to-autosome ratio system like that of Drosophila, and a very few use a ZW system like that of birds and lepidopterans.
.jpg?revision=1)
But the vast majority of angiosperms have perfect flowers; that is containing both male and female sex organs. So how do they avoid self-fertilization? Many plants have self-incompatibility genes to prevent successful pollination between two of their own gametes. However, there is also a morphological solution: heteromorphic flowers. The flowers are perfect but come in two structural types; for example
- long stamens with a short style
- short stamens with a long style
A single plant has one type or the other. If the pollinator has a short tongue, pollination is favored from the first type to the second - but not the reverse. Heteromorphic flowers are not common, and even in the angiosperm families that favor them (e.g., primroses, flax), the same biochemical mechanisms of self-incompatibility that we will find in homomorphic flowers are usually present as well.
Fusion Within and Between Whorls
Determining the merosity of flowers is complicated by floral fusion. In many flowers, parts of a single floral whorl will be partially or completely fused together. When the fusion is between parts of the same whorl, such as the petals fusing together to form a tubular structure (a sympetalous flower), it is called connation. When there is fusion of parts between whorls, such as the stamen fusing the the petals, it is called adnation.
A frequent form of connation occurs within the gynoecium. In an apocarpous gynoecium, the carpels are free. In a syncarpous gynoecium, some or all parts of the carpels are fused (see Figure \(\PageIndex{10}\))



Floral Symmetry
Flowers that have multiple lines of symmetry (like a starfish) are radially symmetrical, also called actinomorphic or regular. Flowers with only a single line of symmetry (like you) are bilaterally symmetrical, also called zygomorphic or irregular (Figure \(\PageIndex{11}\)).

Ovary Position
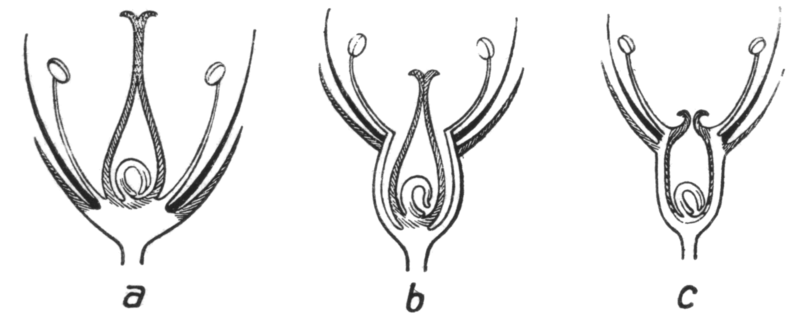
We can use the location of the ovary to further distinguish between flowers (Figure \(\PageIndex{12}\)). If the other whorls of the flower meet below the ovary (the ovary or ovaries look a bit like an egg or eggs in a nest; Figure \(\PageIndex{13}\)), the ovary is superior (on top of the rest of the flower). This means that the rest of the flower parts are below the gynoecium, so we can also call this flower hypogynous (below the gynoecium). The two terms both describe the same situation, but superior refers only to the ovary while hypogynous refers to the flower, in general.
In the opposite situation, the other floral whorls join at a point above the ovary. In this case, the ovary is inferior and the flower is epigynous (on top of the gynoecium). As always, there are less clear situations. In some flowers, as in the rose family, the floral whorls join together and fuse at a point above the ovary, then travel down, around, and below the ovary as a fused unit. This fused unit is called a hypanthium. The ovary is termed semi-inferior, as it is located below the unfused parts of the floral whorls. Because the floral whorls travel around the ovary as the hypanthium, the flower is perigynous (peri- meaning around).

Inflorescences
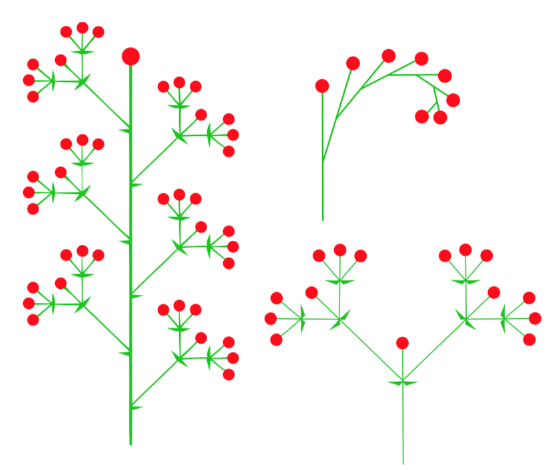
An inflorescence is when, in the place of a single flower, multiple florets are formed. Florets can be sessile or attached by a stem called a pedicel. When leaf-like structures are found within the inflorescence, they are called bracts. There are a wide range of possibilities for the structure and development of inflorescences, though most can be split into four models: raceme-based, cymose, panicle, and intercalate (Figure \(\PageIndex{14}\)). Two models are most widespread.
Raceme-based inflorescences are mostly monopodial, having indeterminate growth. This means that the terminal bud continues producing lateral florets, never truly forming a terminal floret. The inflorescence can be simple or compound (Figure \(\PageIndex{15}\)).
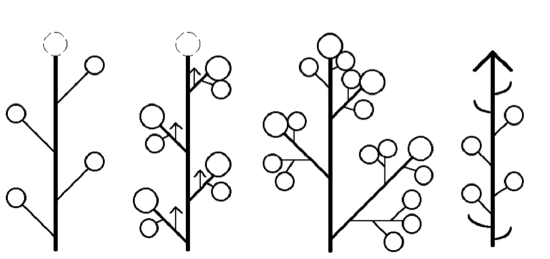
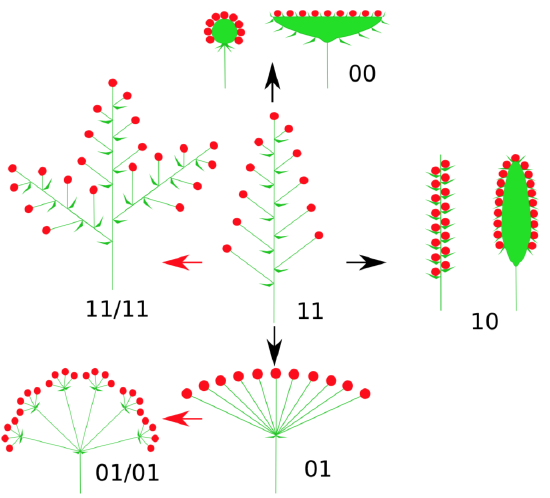
Cymose inflorescences are sympodial, having determinate growth. This means there is a terminal floret that forms first, then other florets are produced laterally (Figure \(\PageIndex{16}\)). The order of lateral floret maturation can be a useful identification feature.
Attribution
Curated and authored by Maria Morrow, CC BY-NC, using the following sources:
- 16.3E Self incompatibility – How Plants Avoid Inbreeding, 16.3D Angiosperm Life Cycle, and 19.1.5 Diversity and Evolutionary Relationships of the Plants from Biology by John. W. Kimball (licensed CC-BY)
- 26.2 Gymnosperms and 32.1 Reproductive Development and Structure from Biology 2e by OpenStax (licensed CC-BY). Access for free at openstax.org.
- 8.2 The Flower and the Fruit from Introduction to Botany by Alexey Shipunov (public domain)
Figure \(\PageIndex{8}\) caption by Melissa Ha.