
2.5.3.2: Polypodiopsida
- Page ID
- 37013

Learning Objectives
- Differentiate between ferns, horsetails, and lycophytes.
- Identify features of vegetative and reproductive shoots of Equisetum.
- Identify features and phases of the fern life cycle; know their ploidy.
- Label a fern gametophyte and sporophyte.
The Polypodiopsida includes the horsetails and ferns. These plants produce leaves with branching veins of vascular tissue called megaphylls. Like the lycophytes, asexual reproduction of the sporophyte can be accomplished via rhizomes. Spores produced in this group develop into gametophytes that can produce both antheridia and archegonia.
Characteristics
- Megaphylls. Leaves have branching veins of vascular tissue. Megaphylls are thought to have evolved from branching stems. Webbing filled in the spaces between the branches, forming the flat blade of the leaf. The arrangement of the leaf veins (vascular bundles) reflects the original branching pattern of the stems.
- Rhizomes. Asexual propogation of the sporophyte through underground stems.
- Homosporous. Haploid spores grow into bisexual gametophytes that produce both antheridia and archegonia, or are capable of producing one or the other, dependent upon conditions.
Equisteum (subclass Equisetidae)
Horsetails are a small group with a single extant genus, Equisetum, which has about 30 different herbaceous species that typically live in moist habitats. The common name comes from the characteristic pattern of branching: whorls or rings of branches arising from an above-ground shoot. The leaves of these plants have been reduced to scales, and instead the segmented stems are photosynthetic. If you look closely at the nodes of a green vegetative shoot, you will see that branches and leaves have not only switched roles, they have also switched places, with the photosynthetic branches emerging below the papery, non-photosynthetic leaves.
Horsetails often grow in sandy places and incorporate silica in their stem epidermis, which gives it an abrasive surface. Because of this, American pioneers would use this plant to scour pots and pans. This is how it received the nickname “scouring rush.” The stem has multiple canals, an analogous characteristic to stems of grasses. The sporangia are associated with hexangular stalked sporangiophores produced on terminal strobili. Within the sporangia, there are elaters that are not separate cells but parts of the spore walls. Gametophytes are typically minute and dioecious, but the plants themselves are homosporous: smaller suppressed gametophytes develop only antheridia while larger gametophytes develop only archegonia.
Gametophyte Morphology
Horsetail gametophytes are reduced and thalloid (figure \(\PageIndex{1}\)). Gametophytes grow from homospores and can produce both antheridia and archegonia.
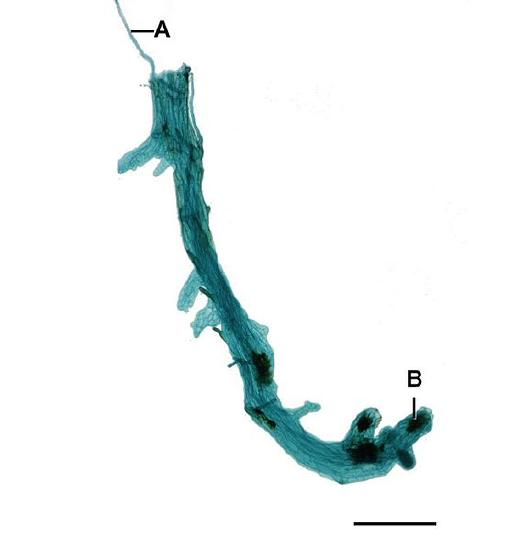
Sporophyte Morphology
In some Equisetum species, there are two different types of shoots produced by the sporophyte: vegetative shoots that perform photosynthesis and reproductive shoots that form strobili and undergo meiosis. In other species, the strobilus is formed at the apex of a photosynthetic shoot.
Vegetative Shoots
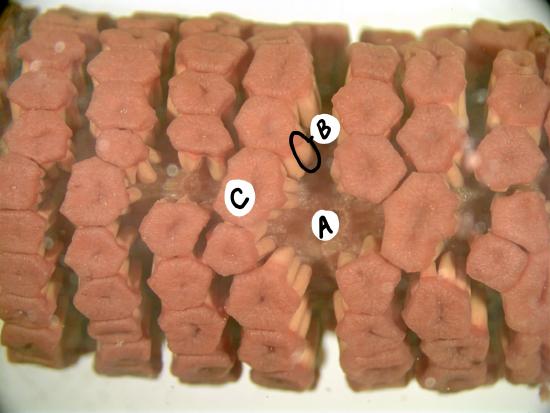
On the vegetative shoot, the leaves are dark, papery and non-photosynthetic. Branches are photosynthetic and produced in whorls. Branches and leaves emerge at nodes, separated by regions of the main stem called internodes. Unlike most plants, the branches emerge below the leaves in the node (Figure \(\PageIndex{2-3}\)). The epidermis of the stems contain silica, which has an abrasive texture.


Reproductive Shoots
Sporangia are produced in a terminal strobilus on the reproductive shoot (figure \(\PageIndex{4-5}\)). In some species, this reproductive shoot lacks chlorophyll and is instead fed through the rhizome of connected vegetative shoots. Spores are photosynthetic and have four hygroscopic arms called elaters.

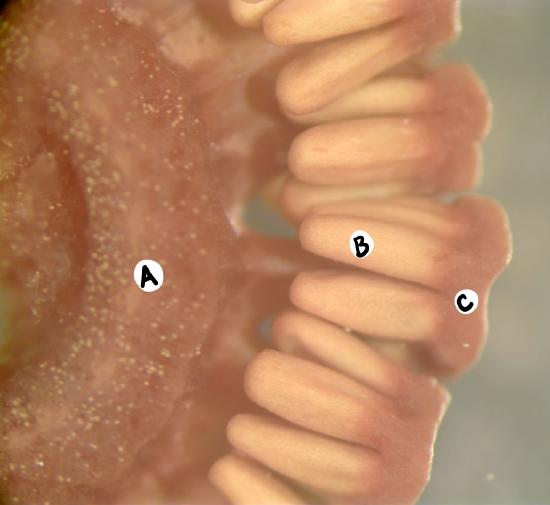
Video \(\PageIndex{1}\): This video shows how the elaters of Equisetum spores respond to changes in humidity. Retrieved from YouTube.
Ferns (subclass Polypodiidae)
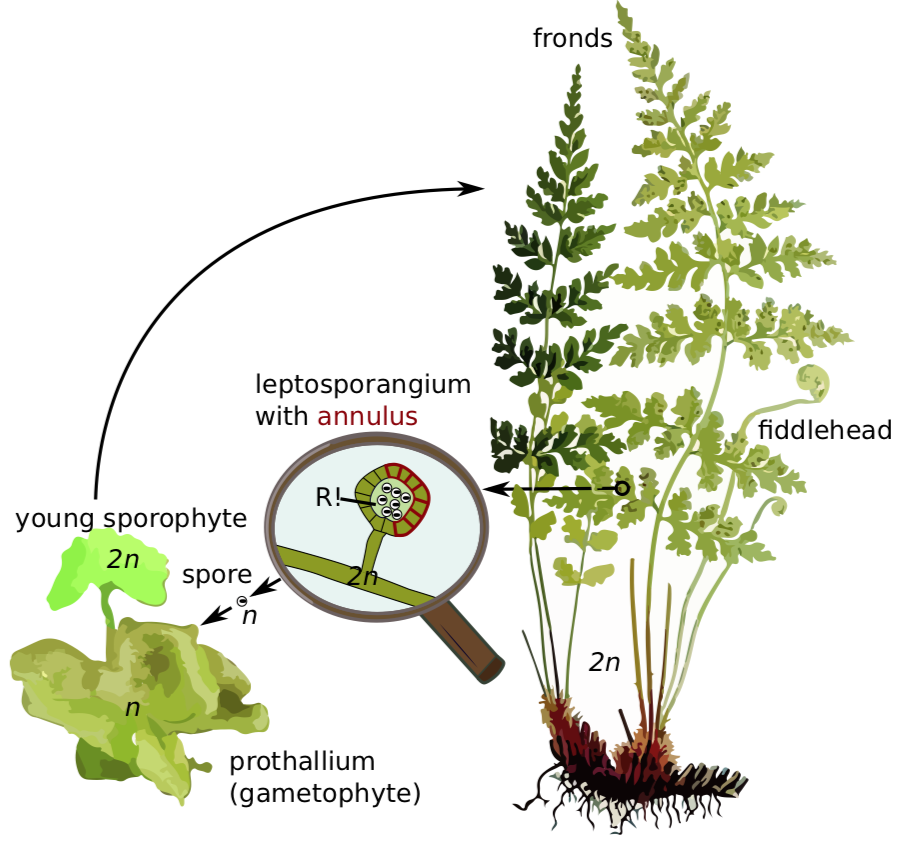
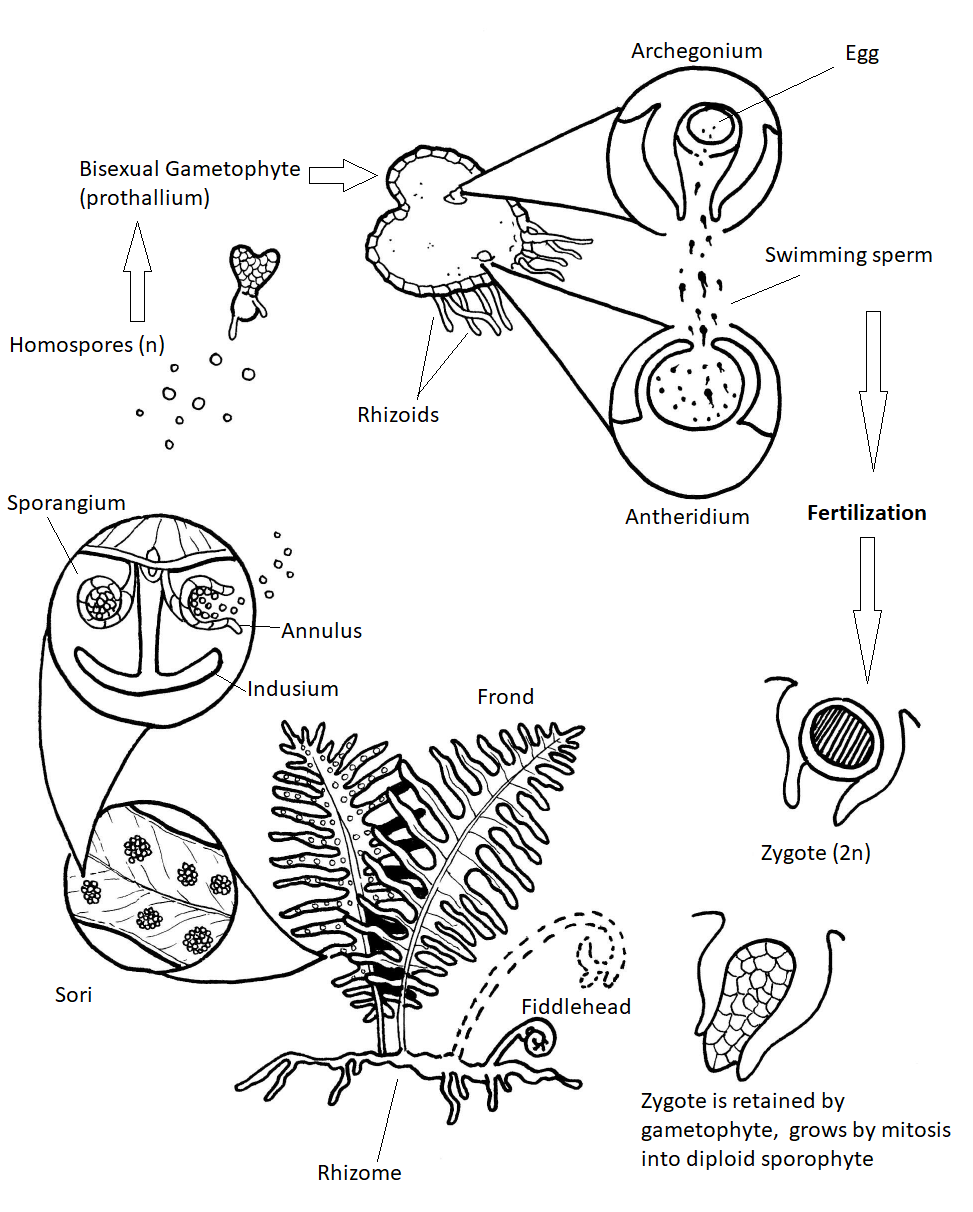
Some 15,000 species of ferns live on earth today. Many of these are found in the tropics where some — the "tree ferns" — may grow to heights of 40 ft (13 m) or more. The ferns of temperate regions are smaller. They are usually found in damp, shady locations. They produce perennial rhizomes that can overwinter. Their leaves, called fronds due to apical growth, emerge from the rhizome each spring as coiled fiddleheads (Figure \(\PageIndex{6}\)). These fiddleheads open through a process called circinate vernation, where the growing tissues are protected at the center of the coil and emerge last.
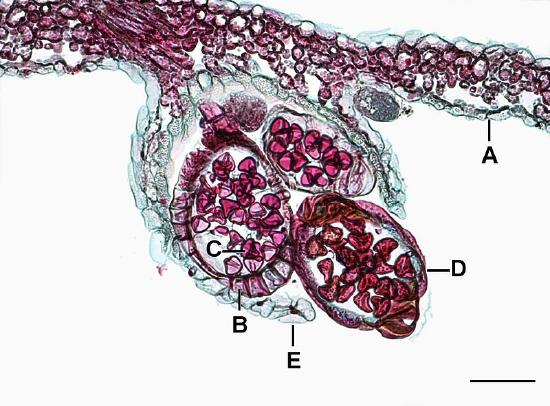
True ferns are megaphyllous: their leaves originated from flattened branches and have branching veins of vascular tissue. True ferns have unique sporangia: leptosporangia (Figure \(\PageIndex{6}\)). Leptosporangia originate from a single cell in a leaf, they have long, thin stalks, and the wall of one cell layer. They also open actively: when sporangium matures (dries), a row of cells with thickened walls on the outside of the sporangium (called an annulus) will shrink slower than surrounding cells and finally would break and release all spores at once. Leptosporangia are grouped in clusters called sori which are often covered with umbrella- or pocket-like indusia. Gametophytes are minute and grow aboveground. While most ferns are homosporous, some genera of true ferns (like the water fern Azolla, water shamrock Marsilea and several others) are heterosporous (Figure \(\PageIndex{7}\)).

True ferns are highly competitive even to angiosperms. In spite of their “primitive” life cycle, they have multiple advantages: abilities to photosynthesize in deep shade (they are not obliged to grow fast), to survive high humidity, and to make billions of reproductive units (spores). Ferns do not need to spend their resources on flowers and fruits, and are also less vulnerable to vertebrate herbivores and insect pests, probably because they do not employ them as pollinators and, therefore, can poison tissues against all animals.
Gametophyte Morphology
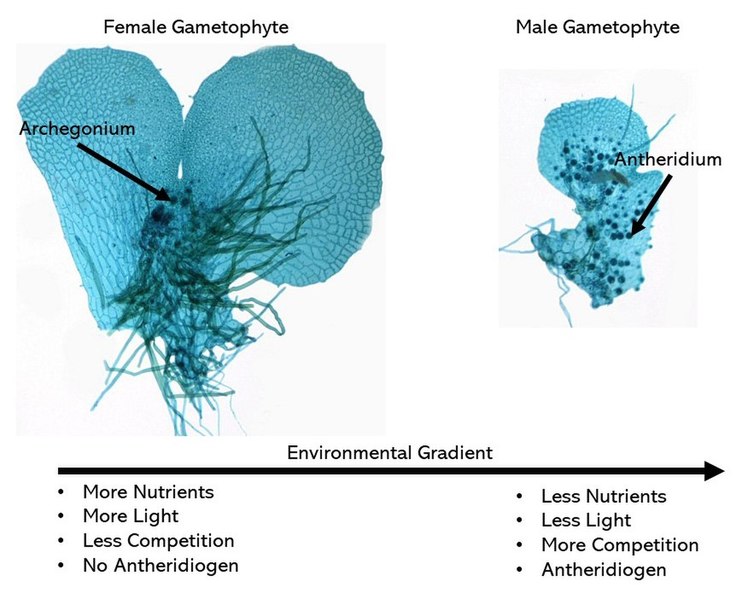
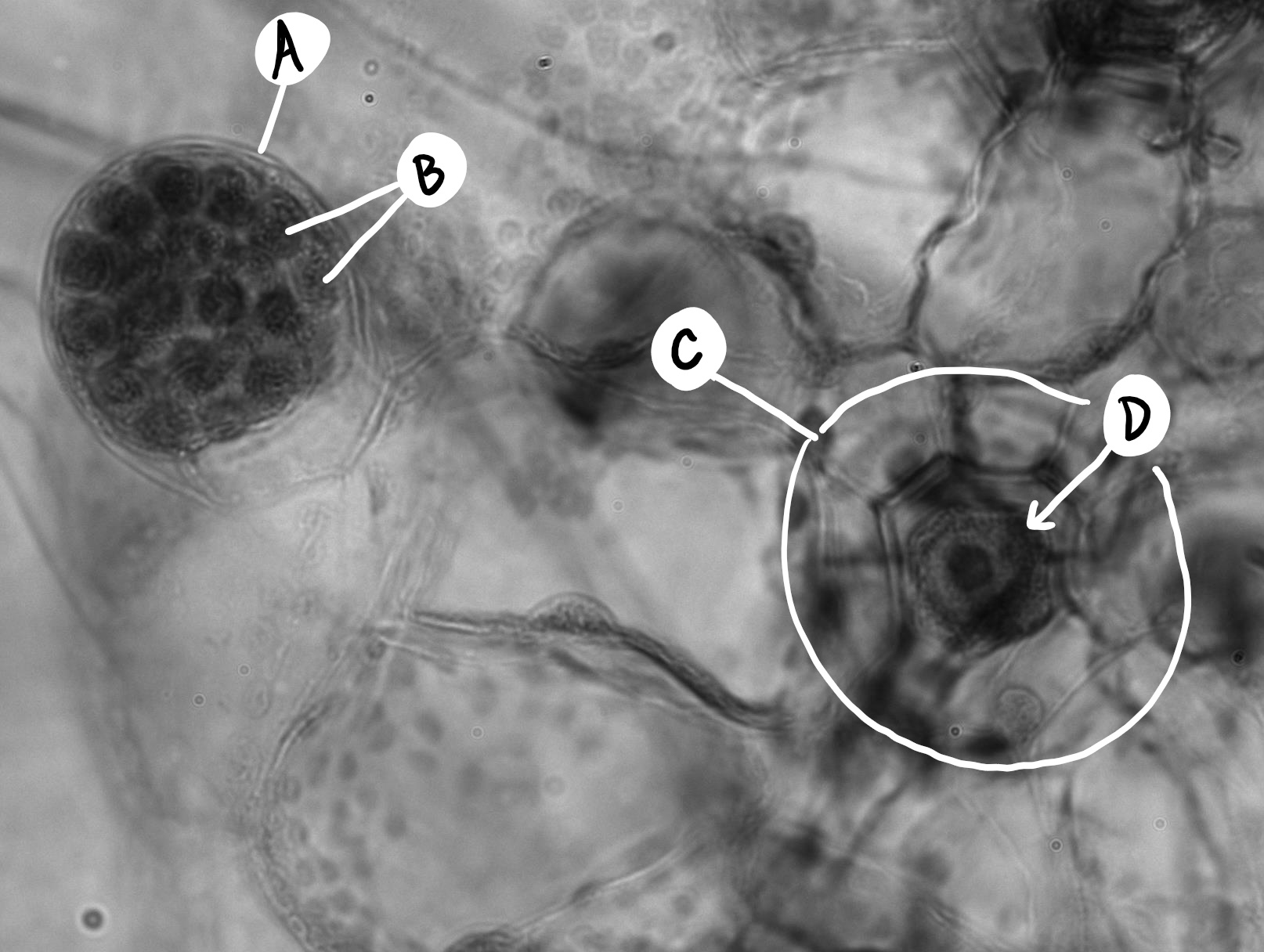
Fern gametophytes are reduced, thalloid, and heart-shaped (Figure \(\PageIndex{8}\)). They are often referred to as a prothallus or prothallium. Rhizoids are produced from the underside of the thallus, just like in the bryophytes. Similar to the horsetails, whether a gametophyte produces antheridia or archegonia can be regulated by environmental cues (Figure \(\PageIndex{9}\)). Each antheridium produces many swimming sperm, but archegonia produce only a single egg (Figure \(\PageIndex{10}\)).
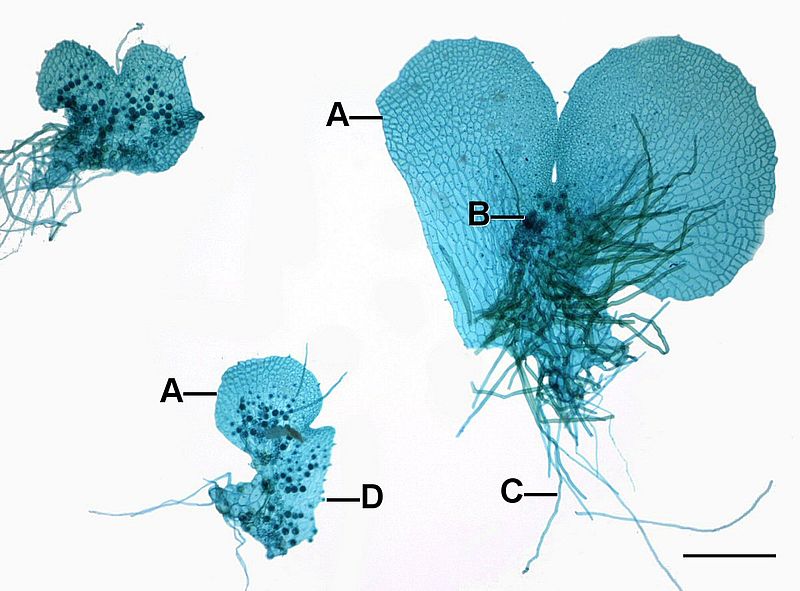
Sporophyte Morphology
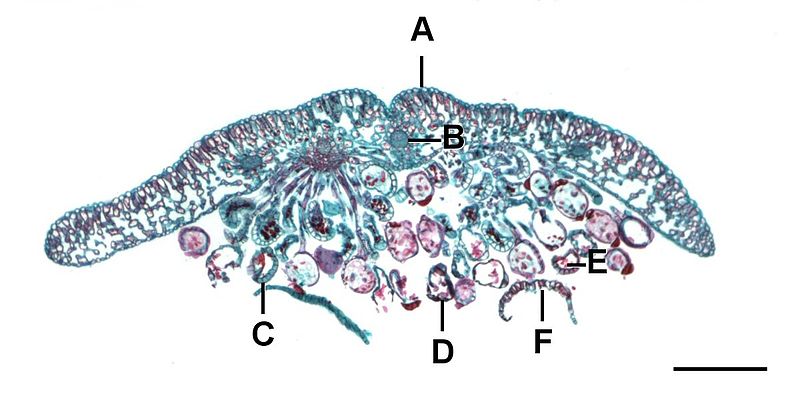
Fern sporophytes are composed of megaphylls, often pinnately compound fronds that emerging as coiled fiddleheads in the spring. Sporangia are produced in clusters called sori (sorus, singular) on the fronds (Figure \(\PageIndex{11}\)).

Circinate vernation is a term used to describe the development of the fern fiddlehead (Figure \(\PageIndex{12}\)) into a frond. Because plants grow apically, it is important to protect the apical meristems in growing organs (as we have seen in both axillary and terminal buds with the protective bud scales). The fiddlehead is essentially a structure that tucks away the growing tips of the fronds. As the frond develops, it gradually unfurls, releasing the tips last.


A sorus (plural, sori) is a cluster of sporangia, often protected by an umbrella-like structure called the indusium as the spores mature (Figure \(\PageIndex{13}\)). Some sori are protected by an extension of the leaf called a false indusium (Figure \(\PageIndex{14}\)), while others lack any protective covering. Each sporangium is lined by an inflated strip of cells called an annulus. When the spores have matured, the cells in the annulus begin to dry out, causing the cells to collapse and pull the sporangium open, releasing the spores.
Full Life Cycle Diagram
Ferns rely on water for dispersal of the sperm, which must swim into an archegonium to fertilize an egg (Figure \(\PageIndex{15}\)). If moisture is plentiful, the sperm swim to archegonia - usually on another prothallus because the two kinds of sex organs generally do not mature at the same time on a single prothallus.
Another method for promoting cross-fertilization: The first spores to germinate develop into prothallia with archegonia. These prothallia secrete a gibberellin into their surroundings. This is absorbed by younger prothallia and causes them to produce antheridia exclusively.
Fertilization restores the diploid number and begins a new sporophyte generation. The embryo sporophyte develops a foot that penetrates the tissue of the prothallus and enables the sporophyte to secure nourishment until it becomes self-sufficient. Although it is tiny, the haploid fern prothallus is a fully-independent, autotrophic plant. Soon, the sporophyte is nutritionally independent. It is the larger, longer-lived stage of the life cycle. To reproduce, many sori are formed on the undersides of the fronds. Within each sporangium of the sorus, the spore mother cells undergo meiosis producing four haploid spores each.
When the humidity drops, the thin-walled lip cells of each sporangium separate, the annulus slowly straightens out, then the annulus snaps forward expelling the spores. Each of these homospores can then grow into a gametophyte capable of producing antheridia and archegonia.
Attributions
- Curated and authored by Maria Morrow using 16.3C Fern Life Cycle and 19.1.5 Diversity and Evolutionary Relationships of the Plants Biology by John. W. Kimball (licensed CC-BY)
- Megaphyll evolution and Salvinia natans information added by Melissa Ha