
28.10: Integrins- Bidirectional Cell Adhesion Receptors
- Page ID
- 15118


\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Learning Goals:
-
Describe Integrin Structure and Composition
- Explain the heterodimeric nature of integrins, including the distinct roles and structural features of α and β subunits.
- Identify key extracellular, transmembrane, and intracellular domains and their functions in ligand binding and intracellular signaling.
-
Understand Ligand Specificity and Integrin Classification
- Classify integrins based on their ligand-binding specificities (e.g., collagen-binding, RGD-binding, laminin-binding, and leukocyte integrins).
- Explain how different αβ pairings determine the specificity and affinity of integrins for various extracellular matrix proteins.
-
Explain Integrin Conformational Changes and Activation Mechanisms
- Compare the “bent-closed” (inactive) and “extended-open” (active) conformations of integrins and discuss the structural changes that occur during activation.
- Describe the processes of “inside-out” and “outside-in” signaling and how they modulate integrin activation and subsequent intracellular signal transduction.
-
Elucidate the Role of Cytoplasmic Interactions in Integrin Function
- Discuss how cytoplasmic proteins such as talin and kindlin bind to the integrin β subunit tails to trigger inside-out signaling and increase ligand affinity.
- Explain the importance of intracellular adaptor proteins in linking integrin activation to downstream signaling pathways and cytoskeletal rearrangements.
-
Integrate Integrin Function with Cellular Processes
- Describe how integrin-mediated adhesion influences cell survival, proliferation, migration, and apoptosis.
- Evaluate the roles of integrins in critical physiological processes, including inflammation, infection, and angiogenesis.
-
Apply Knowledge of Integrin Signaling to Therapeutic Contexts
- Identify how alterations in integrin function and expression contribute to disease processes such as chronic inflammation and cancer.
- Discuss current advances and strategies in integrin-targeted therapies, including their potential for modulating cell adhesion and signaling in pathological conditions.
These learning goals will help you understand the complex structure–function relationships of integrins and their pivotal roles in cellular communication and disease.
This section was derived almost completely, with some modifications and additions, from the following source: Mezu-Ndubuisi, O.J., Maheshwari, A. The role of integrins in inflammation and angiogenesis. Pediatr Res 89, 1619–1626 (2021). https://doi.org/10.1038/s41390-020-01177-9. https://doi.org/10.1038/s41390-020-01177-9. Creative Commons Attribution 4.0 International License. http://creativecommons.org/licenses/by/4.0/. Consult the original article for specific references.
Introduction
Integrins link the extracellular matrix to the intracellular cytoskeleton through a single transmembrane alpha-helical segment. They work with growth factor receptors to regulate cell survival, migration, and division. They contain a very large extracellular domain comprising most of the protein and a very small intracellular domain.
Integrins are heterodimeric transmembrane cell adhesion molecules of alpha (α) and beta (β) subunits arranged in numerous dimeric pairings. These complexes have varying affinities to extracellular ligands. Integrins regulate cellular growth, proliferation, migration, signaling, and cytokine activation and release, playing essential roles in cell proliferation and migration, apoptosis, and tissue repair, as well as in all processes critical to inflammation, infection, and angiogenesis.
Integrins are a family of ubiquitous αβ heterodimeric receptors that exist in multiple conformations and interact with diverse ligands. These molecules mediate interactions between cells and the extracellular matrix (ECM) and thereby serve a critical role in signaling and homeostasis. By facilitating dynamic linkages between the intracellular actin cytoskeleton and the ECM, integrins also transduce external and internal mechanochemical cues and bidirectional signaling across the plasma membrane. Integrins are involved in diverse body processes, including cellular survival, inflammation, immunity, infection, thrombosis, angiogenesis, and malignancy. In this review, we highlight the structure and function of integrins; the mechanisms involved in integrin activation and signaling; their role in inflammation, infection, and angiogenesis; and discuss current advances in integrin-targeted therapies. Understanding the factors that regulate integrin structure, function, and signaling would enable the identification of new therapeutic targets.
Structure of integrins
In mammals, the family of integrins comprises 24 αβ pairs of heterodimeric transmembrane adhesion receptors and cell-surface proteins. These pairings involve 18 α and 8 β subunits as shown in Figure \(\PageIndex{1}\).

With respect to ligand specificity, integrins are generally classified as collagen-binding integrins (α1β1, α2β1, α10β1, and α11β1), Arg-Gly-Asp (RGD)-recognizing integrins (α5β1, αVβ1, αVβ3, αVβ5, αVβ6, αVβ8, and αIIbβ3), laminin-binding integrins (α3β1, α6β1, α7β1, and α6β4), and leukocyte integrins (αLβ2, αMβ2, αXβ2, and αDβ2).
The right-hand side of Figure \(\PageIndex{1}\) shows that the β2 integrin subunit (CD18) can pair with one of the four α subunits (αL-CD11a, αM-CD11b, αX-CD11c, and αD-CD11d), forming leukocyte function-associated antigen-1, Mac1/CR3 (macrophage-1 antigen, complement receptor 3), 150.95/CR4 (complement receptor 4), and CD18/CD11d, respectively.
The structure of the heterodimers and their non-covalent associations are shown in Figure \(\PageIndex{2}\).

Each subunit comprises one large multi-domain extracellular segment, one transmembrane helix, and a short cytoplasmic tail. The extracellular region interacts with extracellular matrix (ECM) ligands and contains about 1104 residues in the α subunit and 778 residues in the β subunit, and shorter cytoplasmic domains with 30–50 residues. The short cytoplasmic tails comprise 20–70 amino acids and mediate interactions with intracellular cytoskeletal and signaling proteins.
In response to intracellular or extracellular stimuli, integrin activation occurs by ligand binding or by changes in the cytoplasmic domains, resulting in elongation and separation of the legs. Integrins appear in a closed or “bent” conformation on resting cells and display a low binding affinity for ligands, rendering them inactive to ligand binding or signal transduction. In contrast, once activated, the integrin shape extends to an open conformation, leading to a high affinity. In a closed conformation, integrins show low ligand-binding affinity, partly due to the bend in the center of the α and β subunits, which brings the ligand-binding site within 5 nm of the cell surface. However, when the conformation is open, the two subunits straighten with increased integrin affinity for the ligand. The initial binding of extracellular ligands causes separation of the cytoplasmic domains, allowing interaction with signal transduction and cytoskeletal molecules during outside-in signaling. In contrast, the separation of the cytoplasmic domains by talin and other activators activates the head to enable ligand binding during inside-out signaling.
The αβ pairings of integrin subunits dictate the specificity of the integrin to a particular ligand, modulate the formation of intracellular adhesion complexes, and regulate downstream signaling. Six α (α1–6) and seven β (β1–7) subunits are known to form several unique αβ subunit associations, as shown in Figure \(\PageIndex{1}\). Interestingly, the earliest discovered integrins, lymphocyte function-associated antigen 1 (integrin αLβ2) and macrophage antigen 1 (integrin αMβ2), derive their specificity from specific α subunits, but these share the same β subunit.
Integrin α subunit family
The integrin α subunits carry a 200 amino acid “inserted” domain, the I-domain (αI). The αI domain is an exclusive ligand-binding site when present on an integrin. αI integrins have 13 extracellular domains in 2 subunits, interacting with various ligands. The I-domains are seen in 6 out of the 15 integrin α subunits.
Integrin alpha-1/beta-1 is a receptor for laminin and collagen. It recognizes the proline-hydroxylated sequence G-F-P-G-E-R in collagen. The human α1 subunit (1179 amino acids) has the domain structure shown in Figure \(\PageIndex{3}\):

The green represents the Van Willibrand Factor Type A domain. The middle pinkish domains are FG-GAP extracellular domains, which are repeated up to seven times in alpha integrins. The organgish domain at the very C-terminus is the transmembrane helix domain (1142-1164). This membrane protein is very different from those we have seen before, as it has just a 15-amino-acid C-terminal tail exposed in the cytoplasm.
Figure \(\PageIndex{4}\) shows an interactive iCn3D model of the predicted structure of human Integrin alpha-1 (AlphaFold, P56199).
.png?revision=1&size=bestfit&width=375&height=246)
The color coding is gray spacefill for the C-terminal transmembrane helix, yellow spacefill for the inhibitor binding pocket, and magenta for the collagen-binding site.
Integrin β subunit family
In humans, integrin β subunits have a cytoplasmic tail with <75 amino acids in length, except the β4 tail, which is about 1000 amino acids long (includes four fibronectin type III repeats). The integrin β tails contain one or two NPxY/F motifs (where x represents any amino acid) that recognize protein modules, specifically phosphotyrosine-binding domains, which are involved in various signaling and cytoskeletal proteins at the cytoplasmic face of the plasma membrane. This recognition occurs through the phosphorylation of the tyrosine (Y) residue in the NPxY/F motif. The integrin β subunit family includes β1–7, which bind the α subunits in different combinations. The most frequently seen β subunit integrin heterodimers are β1.
Although β2 integrins show functional overlap, the corresponding α subunit defines its individual functional properties. The β2/CD18 chain has also received attention because of its involvement in several inflammatory receptors, such as αLβ2, lymphocyte function-associated antigen-1 (LFA-1), and the αMβ2, Mac-1, and complement receptor 3 (CR3). In these β2 integrins, the α subunits bind specific ligands such as the intercellular adhesion molecules (ICAMs). The non-I-domain α subunits in other integrins, such as the laminin-binding α3, α6, and α7, and others that recognize the arginine (R), glycine (G), aspartic acid (D) (RGD) motif (αV, α8, α5, and αIIb), are also closely related to each other.
- The α subunit of each integrin is the primary determinant of its extracellular ligand specificity.
- The β chain binds acidic residues in ICAMs and cytoplasmic adapters such as paxillin, talins, and kindlins to facilitate cellular adhesions with the ECM. Integrins interact with the actin cytoskeleton through the talin- and kindlin-binding motifs in the cytoplasmic domains of their β subunits.
Characteristics of specific integrin heterodimers
Integrin αβ heterodimers are divided into four classes (leukocyte, collagen-binding, Arg-Gly-Asp (RGD)-binding, and laminin-binding integrins (as shown in Figure \(\PageIndex{1}\), based on evolutionary associations, ligand specificity, and restricted expression on white blood cells (β2 and β7 integrins).
- Leucocyte integrins have a common β2 chain linked to CD-18 and bind receptors such as ICAM and plasma proteins such as complement components C3b and C4b.
- Collagen-binding integrins have a common β1 chain that binds various α chains in integrins α1β1, α2β1, α10β1, and α11β1. The α2β1 integrin binds its primary ligand, collagen, and chondroadherin, a matrix protein.
- The RGD-binding integrins have a common αV chain or β1 chain. The RGD peptide motif was first discovered in fibronectin but was later found in several other ECM proteins, such as fibronectin, osteopontin, vitronectin, von Willebrand factor (VWF), and laminin.
Among the 24 human integrin subtypes known to date, eight integrin dimers recognize the tripeptide RGD motif within ECM proteins, namely: αVβ1, αVβ3, αVβ5, αVβ6, αVβ8, α5β1, α8β1, and αIIbβ3. Laminin-binding integrins (α3β1, α6β1, α7β1, and α6β4) mediate the adhesion of cells to basement membranes in various tissues. The α4β1, α9β1, and α4β7 integrin family binds fibronectin in an RGD-independent manner.
Figure \(\PageIndex{5}\) shows an interactive iCn3D model of the structure of α6β1 integrin in complex with laminin-511 (7CEC)
.png?revision=1)
The cyan chain is integrin α6 and the magenta chain is integrin β1. The laminin chains are α (brown), β (orange), and γ (red). Note that each of the laminin subunits interacts with the α6β1 integrin. Two carboxylates on the C-terminal region of the laminin γ chain interact with the metal ion-dependent binding sites on the integrin beta subunit and an Asp 189 in the alpha subunit.
Integrin–ligand binding and consequent activation
Integrins have complex structures and functions. They bind numerous extracellular ligands, intracellular signaling molecules, and the cytoskeleton in a bivalent-cation-dependent manner with varying specificities. Integrins also have many states with multiple conformations and affinities.
Mechanism of integrin-ligand binding and conformational states
Integrins bind cell-surface ligands to promote cellular interactions with the ECM and other cells in the transduction of complex signals that modulate many cellular processes, such as adhesion, migration, and differentiation. These soluble, ECM, or cell surface-bound ligands may include growth factors, structural constituents of the ECM, proteases, cytokines, plasma proteins, microbial pathogens, or receptors specific to immune cells. The affinity and avidity of a ligand may change actively by inside-out signaling in specific pathways. Ligand affinity may vary with the strength of interaction and dissociation of a monovalent protein and its ligand, where ligand avidity refers to its ability to form multiple combinations of bonds.
Integrins exist primarily in three conformational states: bent–closed (inactive; the predominant state), extended–closed (active; low affinity or intermediate state), and extended–open (active; high affinity). The affinity of integrins to various inhibitory and stimulatory ligands is modulated by bivalent cations, which induce a range of conformational changes in integrins ranging from a folded, inactive, and low-affinity state to a high-affinity conformation as shown in Figure \(\PageIndex{2}\). These conformational changes in the extracellular domains of integrins modulate both ligand binding and downstream cellular signaling.
Integrin activation
The activation of integrins increases the affinity of these molecules to extracellular ligands. Integrin tail domains play a critical role in these steps, and any genetic mutations in these parts of integrins can disrupt downstream intracellular signaling. Integrin-mediated signaling across cell membranes is typically bidirectional and termed “outside-in” and “inside-out” signaling. When integrins interact with ECM ligands, a conformational change allows adherence to downstream adaptor molecules in the cell-membrane plane. Once clustered, integrins can recruit and activate kinases such as Src family kinases, focal adhesion, and scaffold molecules such as the adaptor protein p130CRK-associated substrate/breast cancer anti-estrogen resistance 1 (p130CAS/BCAR1). These integrin-associated complexes include discrete active and inactive integrin organizations, which can activate unique signaling pathways.
The extracellular domains of integrins are known to undergo a diverse range of conformational changes that alter the ligand-binding domains. In the cytoplasmic tails of integrins, α-helices are seen as heterodimers, and the β-strands often bind intracellular proteins, such as talin or filamin. The cytoplasmic tail may undergo several specific conformational changes to bind a range of other signal transducers.
This section is derived from Mechanobiology. https://www.mbi.nus.edu.sg/mbinfo/ho...rin-activated/. Creative Commons Attribution-NonCommercial 4.0 International License.
Integrin can be activated from two directions: from the inside by the regulated binding of proteins to the cytoplasmic tails, and from the outside by multivalent ligand binding. In either case, talin binding to the integrin β tails is an essential and final common step. Though the two processes are conceptually separate, they are mutually cooperative, i.e, one can lead to the other. Some structural studies done with force application to mimic ligand/intracellular protein suggested that the combined action of these two events favors the transition from the closed, low-affinity to an open, high-affinity conformation of integrin. Activation leads to bidirectional signaling, which is crucial in various anchorage-dependent events such as adhesion, cell spreading, migration, polarity, and organization of the ECM, leading to physiological changes. Figure \(\PageIndex{6}\) shows the different states of the integrin dimer in inside-out and outside-in signaling.
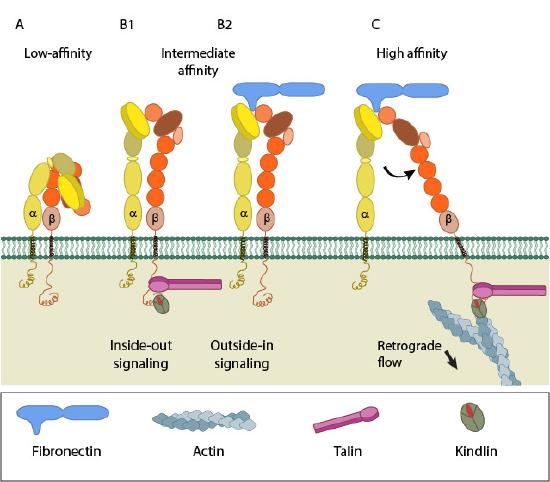
(A) shows integrin in a low-affinity, inactive, bent conformation. (B1) and (B2) show inside-out integrin activation by cytoplasmic proteins or outside-in integrin activation via extracellular matrix (ECM) ligands, both of which lead to the complete extension of the extracellular domains. (C) shows high-affinity and active integrin characterized by separating the cytoplasmic leg domains.
Inside-out signaling
Signals received by other receptors foster the binding of talin and kindlin to the cytoplasmic end of the integrin β subunit at sites of actin polymerization. Substantial information on the signaling pathway leading to activation is available for integrin αIIbβ3.
Talin binds to integrin β-tail via the F3 phospho-tyrosine binding (PTB) domain, a unique interaction with the membrane-proximal (MP) region of the integrin (NPxY motif). This permits competition between a conserved lysine on talin and an aspartic acid on integrin α, essential for α/β salt bridge disruption and sufficient for integrin activation. Additional interactions through the basic patches in the FERM subdomain F2 help to orient the β-subunit to promote spatial separation of the cytoplasmic domains.
Kindlin is also an essential co-activator of integrin and binds to a membrane distal NxxY motif on β-integrin via its FERM F3 subdomain. A preceding threonine patch on integrins β1 and β3 that gets phosphorylated and a tryptophan on kindlin F3 are also required for binding. However, kindlins are not known to activate integrins independently but may render integrin-specific effects.
Outside-in signaling
Ligand binding to the external domain causes conformational changes that increase ligand affinity, modify protein-interaction sites in the cytoplasmic domains, and, hence, the resulting signals.
Besides conformational changes that extend integrin dimers, multivalent ligand binding leads to the clustering of integrins, activating the Src family of kinases (SFKs) by autophosphorylation. SFKs phosphorylate tyrosines of the integrin cytoplasmic domain (NPxY motifs) and other proteins, leading to:
- control of ligand binding strength
- alteration of binding with signaling molecules (kinases, GTPases, and adaptors), which constitute dynamic adhesion structures such as focal adhesions and podosomes
Nevertheless, whether clustering triggers outside-in signaling to facilitate integrin activation or occurs after integrin activation is uncertain.
David G. Menter, Raymond N. DuBois, "Prostaglandins in Cancer Cell Adhesion, Migration, and Invasion", International Journal of Cell Biology, vol. 2012, Article ID 723419, 21 pages, 2012. https://doi.org/10.1155/2012/723419. Creative Commons Attribution License,
Let's look at another, more detailed representation of integrin states. Each αβ dimer recognizes a different intercellular adhesion molecule (ICAM), ligand, or protein substrate in the basement membrane or extracellular matrix. The α subunit dictates the ligand specificity by a seven-bladed β-propeller head domain connected to a leg support structure made of a "thigh", a "calf"-1, a "calf"-2, a transmembrane, and a cytoplasmic domain. The β subunit interacts with the cell cytoskeleton. It contains an N-terminal plexin-semaphorin-integrin (PSI) domain, a hybrid domain, a βI domain, four cysteine-rich epidermal growth factor (EGF) repeats, a transmembrane, and a cytoplasmic domain.
In many cases, the N-terminal β-I domain of a β subunit inserts into the 7-bladed β-propeller domain of an α subunit (α1, α2, α10, α11, αL, αM, αX, and αD) to form a bulbous-binding headpiece complex. The formation of integrin receptor complexes depends on divalent cations (i.e., Ca2+, Mn2+, Mg2+) that bind to metal-ion-dependent adhesion site (MIDAS) motifs in the α subunits and adjacent to MIDAS (ADMIDAS) motifs in β subunits found in the N-terminus of these receptors. Together they joined α and β subunit termini form an N-terminal headpiece. Figure \(\PageIndex{7}\) shows these detailed features of the integrin dimer structure.

Three conformational states exist for α and β subunit complexes. (1) The unliganded conformation has a closed headpiece and a bent receptor structure with the EGF domains of the β-subunit touching the calf-1-calf-2 domains of the α-subunit. (2) The headpiece remains closed, but structural changes in the β-subunit EGF domains cause a separation from the calf-1-calf-2 domains of the α-subunits, causing an extended structure. (3) Conformational changes in the β6-α7 loops expose the ligand-binding site along with a complete separation of the β-subunit from the calf-1-calf-2 domains in the α-subunit. These conformational changes engage the specific integrin headpiece with its ligand.
Figure \(\PageIndex{8}\) shows an interactive iCn3D model of the headpiece of integrin αIIbβ3 in the headpiece extended and open conformation (3FCU)
.png?revision=1&size=bestfit&width=659&height=310)
The αIIb part of the headpiece is colored based on the secondary structure, with the yellow sheets comprising the beta propeller secondary structure motif. The gray chain is the β3 chain. The side chains of the β3 chain forming the binding interface between the αIIb and β3 chains are displayed as colored sticks. The three close metal ions (2 Ca2+ and 1 Mg2+) are important for ligand binding, with the Mg2+ involved in coordination to acidic side chains of integrin ligands. These metal ions are present before ligand binding. The RGD binding motif of some integrin ligands binds to Mg2+ through their aspartate. Without bound Mg2+, acidic side chains around the site would interfere with binding.
It appears that lateral forces are most important in activating integrins. This is in contrast to tensile forces, which act along the length of the receptor. Tensile forces appear to stabilize the closed, extended low-affinity form. In contrast, lateral forces at the beta subunit, a mimic for moving cytoskeletal filaments, stabilized the open, extended, high-affinity form. This links conformational allosteric changes to cytoskeletal changes. The mechanism for activation is hence mechanochemical.
Similar to conventional cell surface signal transducing receptors, integrins bind ligands and transmit information in an “outside-in signaling” as shown in the top panel of Figure \(\PageIndex{9}\). “Outside-in signaling” behavior typically involves the engagement of integrins with the extracellular matrix or ICAM surface receptors. When external factors bind to exposed ligand binding sites on integrins, this results in conformational changes described in the previous section. Most ECM proteins exhibit multivalent or recurrent molecular patterns, which trigger integrin clustering. As cells engage the repetitive patterns in the ECM, these events coincide, activating intracellular signals. The myriad extracellular signals that cells encounter in their microenvironment mediate cell polarity, cytoskeletal structure, adhesion, migration, invasion, gene expression, cell survival, and proliferation.

ECM ligands' “outside-in” binding to cell surface integrins stimulates conformational changes that activate focal adhesion kinase (FAK). FAK is then autophosphorylated on Tyrosine 397 near the catalytic domain, which binds Src. FAK contains a central kinase domain bordered by a FERM (protein 4.1, ezrin, radixin, and moesin homology) domain at the N-terminus and a focal adhesion targeting (FAT) sequence at the C-terminus. Activated Src interacts with human enhancer of filamentation1 (HEF1) and p130 CRK-associated substrate (p130CAS) scaffold proteins that help to regulate Src-FAK-Crk interactions with Rac positively. FAK also activates (PKL/Git2)-β-Pix complexes, and β-pix then serves as an exchange factor for Cdc42 or a scaffold protein to promote signaling via Rac and p21-activated protein kinases (PAK). FAK also interacts with actin-related proteins (ARP2 and ARP3), which are regulated by the Wiskott-Aldrich Syndrome Protein (WASP). ARP2/ARP3 initiates the polymerization of new actin filaments. FAK also influences actin contraction and polarization through another GTPase protein, Rho. The regulation of Rho GTPase hydrolysis of GTP (active) to GDP (inactive) form occurs through the opposing activities of guanine nucleotide exchange factor (GEFs). The GTPase regulator associated with FAK (GRAF) and p190RhoGAP blocks actin cytoskeleton changes.
In contrast, PDZRhoGEF and p190RhoGEF both serve to activate Rho. “Outside-in signaling” transfers integrin-mediated external signals to the inside of cells.“Inside-out signaling” depends on talin and kindlin. Both talin and kindlin contain FERM (4.1/ezrin/radixin/moesin) domains and a highly conserved C-terminal F3 domain. Talins bind β integrin and actin through the C-terminus, and also vinculin. Kindlins bind integrins, the cell membrane, and various actin adaptor proteins like migfilin or integrin-linked kinase (ILK). Talin activation occurs through G-protein-coupled receptors that increase cytoplasmic Ca2+ and diacylglycerol. This activates GEF function with Ras-proximate-1/Ras-related-protein-1-(Rap1-) GTPase. Rap1 then binds to the Rap1-GTP-interacting adaptor molecule (RIAM). RIAM recruits talin to the membrane and the α and β integrin cytoplasmic domains. Kindlin interacts with β integrin cytoplasmic domain stabilizing the activated state of the integrin complex. “Inside-out signaling” strengthens adhesive contacts and the appropriate force necessary for integrin-mediated cell migration, invasion, ECM remodeling, and matrix assembly.
In the case of “outside-in signaling” initiated by ECM proteins, a single ligand-binding event can trigger integrin activation. However, repetitive, regularly spaced molecular patterns provide a more effective stimulus. These adhesion-dependent sensory mechanisms lead to signal transduction inside the cell by activating multiple pathways. Focal adhesions are often formed due to cell interactions with the ECM substrata, which initiate signal transduction via kinase cascades and other mechanisms.
Integrins in inflammation and infection
In the resting state, β2 integrins are expressed specifically on leucocyte receptors. During inflammation, the inflammatory cytokines activate these integrins, promote cellular adherence to counter-receptors such as ICAMs, and promote phagocytosis and cytotoxic killing. Integrin receptors on leukocytes, such as the macrophage-1 antigen (Mac-1, also known as CR3, αMβ2, CD11b/CD18) interact with platelet antigens such as the glycoprotein Ibα (GPIbα) during inflammation. Integrins bind to the pro-domain of transforming growth factor (TGF)-β1 to activate and promote its secretion. The pro-TGF-βs are biosynthesized and stored in tissues in latent forms, and integrins αVβ6 and αVβ8 can uniquely bind and activate pro-TGF-β1 and pro-TGF-β3. The αVβ6 integrin is known to specifically bind the RGDLXXL/I motif in TGF-β1 and TGF-β3.
β2 integrins promote the recruitment of leukocytes to the sites of inflammation by promoting the adhesion of circulating leukocytes to vascular endothelium, transendothelial migration, the formation of immunological synapses in leukocytes, and inflammatory signaling in involved cells. β integrins on the leukocyte surface are also involved in the tethering, rolling, and adhesion of leukocytes to activated endothelial cells. β2 integrins can also initiate intracellular signaling pathways in macrophages and neutrophils and stimulate cytokine secretion from these cells directly or in synergy with Toll-like receptors (TLRs). Integrins may also integrate the impact of the epidermal growth factor receptor, platelet-derived growth factor receptor, insulin receptor, met receptor superfamily (hepatocyte growth factor receptor), and the vascular endothelial growth factor receptor (VEGFR) in inflammatory cells.
β2 integrins are essential adhesion, leukocyte recruitment, and immunological signaling regulators. These integrins mediate adhesive interactions between myeloid cells, endothelial cells, antigen-presenting cells, T cells, and the ECM. L-selectin, the CCR7 chemokine receptor, interacts with specific carbohydrate epitopes on the endothelium and promotes leukocyte rolling and transmigration through the vascular endothelium. Leukocyte rolling induces a rapid, although transient, increase in the affinity of the β1 and β2 integrins to the endothelial ligands. Conformational changes in the structure of the inserted (I) domain of the αL subunit of LFA-1 enhance firm leukocyte adhesion under shear flow.
Summary
Integrins are essential transmembrane receptors that bridge the extracellular matrix (ECM) to the intracellular cytoskeleton, orchestrating a wide range of cellular processes including adhesion, migration, proliferation, and survival. These heterodimeric proteins consist of one α and one β subunit, each contributing distinct structural and functional domains. The large extracellular portion mediates ligand binding—determining specificity for ECM components such as collagen, laminin, and RGD-containing proteins—while the single transmembrane helix anchors the integrin in the plasma membrane, and the short cytoplasmic tail connects to intracellular adaptor proteins and signaling molecules.
Integrins exist in multiple conformational states that dictate their ligand affinity. In the resting, bent–closed conformation, integrins exhibit low affinity for their ligands. Upon activation through either external ligand binding (outside-in signaling) or intracellular interactions (inside-out signaling mediated by proteins like talin and kindlin), integrins extend into an open, high-affinity conformation. This conformational shift not only enhances ECM binding but also triggers downstream signaling cascades, often involving kinases such as Src family members and focal adhesion kinase (FAK), which in turn regulate cytoskeletal dynamics and cellular responses.
Moreover, integrins play pivotal roles in mechanotransduction by transmitting both external mechanical cues and internal signals, thereby influencing processes critical to tissue repair, inflammation, infection, and angiogenesis. Alterations in integrin expression or function are implicated in various pathological conditions, making them attractive targets for therapeutic intervention.
In summary, the study of integrins encompasses their unique structure, diverse ligand specificities, dynamic activation mechanisms, and integral roles in bidirectional signal transduction across the cell membrane—a complex interplay that is crucial for maintaining cellular homeostasis and responding to environmental changes.