
6.4: Enzyme Inhibition
- Page ID
- 102267


\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)(Learning goals written by Claude, Sonnet 4.6, Anthropic)
Reversible Inhibition: Mechanisms and Kinetic Signatures
- Distinguish competitive, uncompetitive, and noncompetitive/mixed inhibition mechanisms by describing where the inhibitor binds (free E only for competitive; ES only for uncompetitive; both E and ES for mixed/noncompetitive), and predict the effect of each mechanism on KMapp and VMapp — using Le Chatelier's principle to rationalize why competitive inhibition increases KMapp while leaving VM unchanged, uncompetitive inhibition decreases both KMapp and VMapp by the same factor (1 + I/Kii), and noncompetitive inhibition decreases VMapp while leaving KM unchanged.
- Interpret Lineweaver-Burk double-reciprocal plots for each inhibition type — recognizing the hallmark patterns of lines intersecting on the y-axis (competitive, same 1/VM), parallel lines with changed intercepts (uncompetitive), and lines intersecting on the x-axis (noncompetitive) — and use slopes and intercepts to extract KIS and/or Kii from experimental data.
- Write and apply the modified Michaelis-Menten velocity equations for each inhibition type: v₀ = VM[S]/(KM(1 + I/KIS) + [S]) for competitive inhibition, where KMapp = KM(1 + I/KIS); v₀ = VM[S]/(KM + [S](1 + I/Kii)) for uncompetitive inhibition, where both KMapp = KM/(1 + I/Kii) and VMapp = VM/(1 + I/Kii); and the mixed equation where both KMapp and VMapp change with inhibitor concentration governed by KIS and Kii independently.
- Explain the concept of the specificity constant kcat/KM in the context of competing substrates — deriving that the ratio of initial velocities for two competing substrates at equal concentration equals the ratio of their kcat/KM values — and use this relationship to explain how an enzyme selects among multiple substrates in vivo.
Special Cases and Physiological Contexts
- Explain product inhibition as a special case of competitive inhibition in which the structurally similar product P occupies the active site and inhibits further turnover of S, describe its effect on progress curves (downward curvature at early time points when product accumulates), and distinguish this from simple reversible product release.
- Contrast in vitro inhibition analysis (fixed enzyme, varying S at several fixed I concentrations, independent variable is [S]) with in vivo inhibition (enzyme embedded in a metabolic pathway with relatively constant flux v, where S concentration is determined by v rather than varied independently) — deriving that under constant flux conditions, competitive inhibition gives a linear relationship between S/KM and I/KIS whereas uncompetitive inhibition causes S/KM to "blow up" to infinity at I = Kii — and explain why this makes uncompetitive inhibitors theoretically more effective at disrupting metabolic pathways in vivo than competitive inhibitors.
- Explain how pH and temperature affect enzyme activity by altering the protonation states of catalytically essential active site residues (affecting kcat), substrate binding (affecting KM), and global protein conformation (affecting both), and identify which amino acid side chains are plausible candidates for the pH-sensitive residues in enzymes whose activity profiles peak at specific pH values.
Irreversible Covalent Inhibition
Given what you already know about protein structure, it should be easy to determine how to inhibit an enzyme. Since structure mediates function, anything that significantly alters an enzyme's structure would inhibit its activity. Hence, extremes of pH and high temperature, both of which can denature the enzyme, would irreversibly inhibit it unless it could refold properly. Alternatively, we could add a small molecule that interacts noncovalently with the enzyme, either altering its conformation or directly preventing substrate binding. Finally, we could covalently modify certain side chains, which, if they are essential to enzymatic activity, would irreversibly inhibit the enzyme.
We previously discussed the types of reagents that could chemically modify specific side chains critical to enzymatic activity. For example, iodoacetamide might abolish enzyme activity if a cysteine side chain is required. However, these reagents will usually modify several side chains. Determining which is critical for the binding or catalytic conversion of the substrate can be difficult. One way would be to protect the active site with saturating concentrations of a reversible-binding ligand. Then, the chemical modification can be performed at varying reaction times. The critical side chain would be protected from chemical modification, with the extent of protection determined by the KD, the concentration of the protecting ligand, and the reaction time.
The rest of the chapter will deal with reversible, noncovalent inhibition
Competitive Inhibition
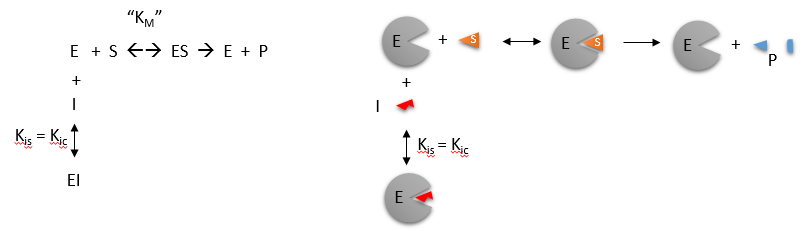
Reversible Competitive inhibition occurs when the substrate (S) and the inhibitor (I) bind to the same site on the enzyme. In effect, they compete for the active site and bind in a mutually exclusive fashion. This is illustrated in the chemical equations and molecular cartoons shown in Figure \(\PageIndex{1}\).
\begin{equation}
v_0=\frac{V_M S}{K_M\left(1+\frac{I}{K is}\right)+S}
\end{equation}
There is another type of inhibition that would give the same kinetic data. If S and I were bound to different sites, and S was bound to E and produced a conformational change in E such that I could not bind (and vice versa), then the binding of S and I would be mutually exclusive. This is called allosteric competitive inhibition. Inhibition studies are usually performed at several fixed, non-saturating I concentrations and varying S concentrations.
The key kinetic parameters to understand are VM and KM. Let us assume for ease of equation derivation that I binds reversibly and with rapid equilibrium to E, with a dissociation constant KIS. The "s" in the subscript "is" indicates that the slope of the 1/v vs. 1/S Lineweaver-Burk plot changes while the y-intercept stays constant. KIS is also named KIC, where the subscript "c" stands for competitive inhibition constant.
A look at the top mechanism shows that even in the presence of I, as S increases to infinity, all E is converted to ES. That is, there is no free E to which I could bind. Now, remember that VM ,= kcatE0. Under these conditions, ES = E0; hence, v = VM. VM is not changed. However, the apparent KM, KMapp, will change. We can use LaChatelier's principle to understand this. If I binds to E alone and not ES, it will shift the equilibrium of E + S → ES to the left. This would increase the KMapp (i.e., it would appear that the affinity of E and S has decreased). The double reciprocal plot (Lineweaver-Burk plot) offers a great way to visualize the inhibition as shown in Figure \(\PageIndex{2}\).
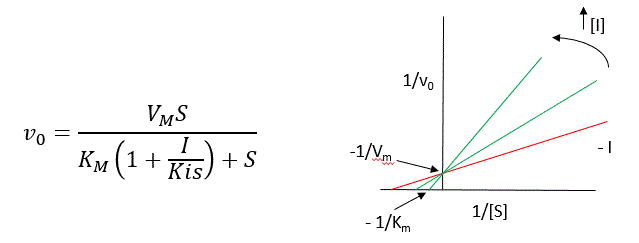
In the presence of I, VM does not change, but KM appears to increase. Therefore, 1/KM, the x-intercept on the plot, will get smaller and closer to 0. Therefore, the plots will consist of a series of lines, with the same y-intercept (1/VM), and the x-intercepts (-1/KM) move closer to 0 as I increases. These intersecting plots are the hallmarks of competitive inhibition.
Here is an interactive graph showing v0 vs [S] for competitive inhibition with Vm and Km both set to 100. Change the sliders for [I] and Kis and see the effect on the graph.
Here is the interactive graph showing 1/v0 vs 1/[S] for competitive inhibition, with Vm and Km both set to 10.
Note that in the first three inhibition models discussed in this section, the Lineweaver-Burk plots are linear in the presence and absence of an inhibitor. This suggests that plots of v vs. S in each case would be hyperbolic and conform to the Michaelis-Menten equation, with potentially different apparent VM and KM values.
An equation for v0 in the presence of a competitive inhibitor is shown in the above figure. The only change from the equation for the initial velocity in the absence of the inhibitor is that the KM term is multiplied by 1 + I/Kis. Hence KMapp = KM(1+I/Kis). This shows that the apparent KM does increase as we predicted. KIS is the inhibitor dissociation constant. The apparent KM changes with increasing I, which affects the slope of the double reciprocal plot.
If the data were plotted as v0 vs. log S, the plots would be sigmoidal, as we saw for plots of ML vs. log L in Chapter 5B. In the case of a competitive inhibitor, the plot of v0 vs log S in the presence of different fixed concentrations of inhibitor would consist of a series of sigmoidal curves, each with the same VM, but with different apparent KM values (where KMapp = KM(1+I/Kis), progressively shifted to the right. Enzyme kinetic data is rarely plotted this way. These plots are mostly used for simple binding data for the M + L ↔ ML equilibrium in the presence of different inhibitor concentrations.
Reconsider our discussion of the simple binding equilibrium, M + L ↔ ML. For fractional saturation Y vs a log L graphs, we considered three examples:
- L = 0.01 KD (i.e., L << KD), which implies that KD = 100L. Then Y = L/[KD+L] = L/[100L + L] ≈ 1/100. This implies that irrespective of the actual [L], if L = 0.01 KD, then Y ≈0.01.
- L = 100 KD (i.e,. L >> KD), which implies that KD = L/100. Then Y = L/[KD+L] = L/[(L/100) + L] = 100L/101L ≈ 1. This implies that irrespective of the actual [L], if L = 100 Kd, then Y ≈1.
- L = KD, then Y = 0.5
These scenarios show that if L varies over four orders of magnitude (0.01KD < KD < 100KD), or, in log terms, from
-2 + log KD < log KD< 2 + log KD), irrespective of the magnitude of the KD, that Y varies from approximately 0 to 1.
In other words, Y varies from 0 to 1 when L varies from log KD by +2. Hence, plots of Y vs log L for a series of binding reactions of increasingly higher KD (lower affinity) would reveal a series of identical sigmoidal curves shifted progressively to the right, as shown below in Figure \(\PageIndex{3}\).

The same would be true of v0 vs. S in the presence of different concentrations of a competitive inhibitor, for initial flux (Jo) vs ligand outside, or ML vs. L (or Y vs L).
In many ways, plots of v0 vs. lnS are easier to visually interpret than plots of v0 vs. S. As noted for simple binding plots, textbook illustrations of hyperbolas are often misdrawn, showing curves that level off too quickly as a function of [S] as compared to plots of v0 vs lnS, in which it is easy to see if saturation has been achieved. In addition, as the curves above show, multiple complete plots of v0 vs lnS at varying fixed inhibitor concentrations or for variant enzyme forms (different isoforms, site-specific mutants) over a broad range of lnS can be generated, facilitating comparisons of experimental kinetics under these conditions. This is especially true if Km values differ widely.
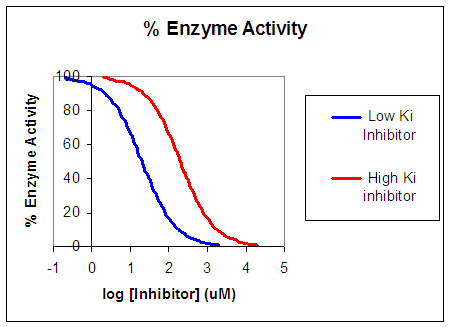
Now that you are more familiar with binding and enzyme kinetics curves, in the presence and absence of inhibitors, you should be able to apply the above analysis to inhibition curves where the binding or the initial velocity is plotted at varying competitive inhibitor concentrations at different fixed nonsaturating concentrations of ligand or substrate. Consider the activity of an enzyme. Let's say that at some reasonable substrate concentration (not infinite), the enzyme is approximately 100% active. If a competitive inhibitor is added, the enzyme's activity decreases until, at saturating (infinite) I, no activity remains. Graphs showing this are shown below in Figure \(\PageIndex{4}\).
Progress Curves for Competitive Inhibition
In the previous section, we explored how important progress curve (Product vs time) analyses are in understanding both uncatalyzed and enzyme-catalyzed reactions. We are aware of no textbooks that cover progress curves for enzyme inhibition. Yet, progress curves are what most investigators record and analyze to determine initial rates (v0) and to calculate VM, KM, and inhibition constants, as described above. We will use Vcell to produce progress curves for reversibly inhibited enzyme-catalyzed reactions.
 MODEL
MODEL
Competitive Inhibition with constant [I]:
No inhibition (left) and competitive inhibition (right)
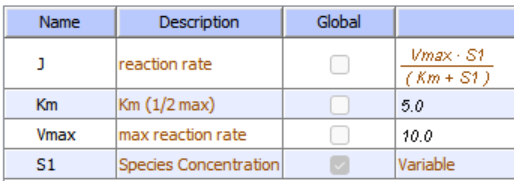
Initial conditions for no inhibition
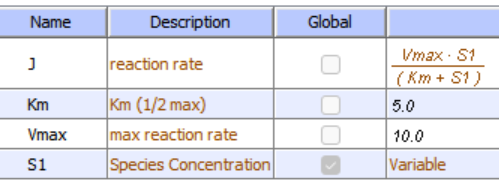
Initial conditions for competitive inhibition
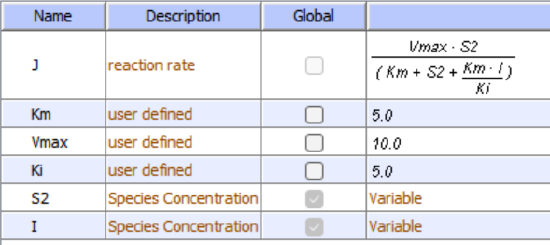
I is fixed for each simulation (as it is not converted to a product) but can be changed in the simulation below.
Select Load [model name] below
Select Start to begin the simulation.
Select Plot to change Y axis min/max, then Reset and Play | Select Slider to change which constants are displayed. For this model, select Vm, Km, Ki and I | Select About for software information.
Move the sliders to change the constants and see changes in the displayed graph in real-time.
Time course model made using Virtual Cell (Vcell), The Center for Cell Analysis & Modeling, at UConn Health. Funded by NIH/NIGMS (R24 GM137787); Web simulation software (miniSidewinder) from Bartholomew Jardine and Herbert M. Sauro, University of Washington. Funded by NIH/NIGMS (RO1-GM123032-04)
The graphs from your initial run show the concentrations of S, P, and I as a function of time for just the initial conditions shown above. In typical initial rate analyses of competitive inhibition, at least three sets of reactions are run with the same varying substrate concentrations and different fixed inhibitor concentrations. In the analyses above, [I] is fixed at 5 uM.
Conduct a series of runs at different values of I. Vary the KI, the dissociation constant for the EI complex, as follows:
- I << KI, the dissociation constant for the EI complex
- I >> KI, the dissociation constant for the EI complex. Then, download the data and determine the initial rate for each of the initial conditions.
Figure \(\PageIndex{5}\) shows an interactive iCn3D model of human low molecular weight phosphotyrosyl phosphatase bound to a competitive inhibitor (5PNT)
.png?revision=1&size=bestfit&width=390&height=330)
 Figure \(\PageIndex{5}\): Human low molecular weight phosphotyrosyl phosphatase bound to a competitive inhibitor (5PNT). (Copyright; author via source).
Figure \(\PageIndex{5}\): Human low molecular weight phosphotyrosyl phosphatase bound to a competitive inhibitor (5PNT). (Copyright; author via source).
Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...XsEacG2tixDDi9
The competitive inhibitor, the deprotonated form of 2-(N-morpholino)-ethanesulfonic acid (MES), is actually the conjugate base of the weak acid (pKa = 6.15) of a commonly used component of a buffered solution. It is shown in color sticks with the negatively charged sulfonate at the bottom of the active site pocket. The amino acids comprising the active site binding pocket are shown as color sticks underneath the transparent colored surface of the binding pocket. The normal substrates for the enzyme are proteins phosphorylated on tyrosine side chains, so the sulfonate mimics the negatively charged phosphate group of the phosphoprotein target.
Two special cases of competition inhibition
Product Inhibition
Let's look at an enzyme that converts reactant S to product P. Since P arises from S, they may have structural similarities. For example, what if GTP is the reactant and GDP the product? If so, then P might also bind in the active site and inhibit the conversion of S to P. This is called product inhibition. It probably occurs in most enzymes, and when it does, it will begin bending downward at the beginning of the progress curve for P formation. If the product binds very tightly, it may lead to a significant underestimation of the enzyme's initial velocity (v0) or flux (J0). Let's use Vcell to explore product inhibition. The model will explore two reactions:
- E + R ↔ ER → E + Q (no product inhibition)
- E + S ↔ ES → E + P (with product inhibition)
The chemical equation above does not explicitly show the product P binding the enzyme to form an EP complex. An actual reaction diagram showing the inhibition of an enzyme by an inhibitor I and by the product P is shown in Figure \(\PageIndex{6}\) below.

Figure \(\PageIndex{6}\): reaction diagram showing inhibition of an enzyme by an inhibitor I and by the product P
Vcell uses much simpler diagrams because it is most often used to model entire pathways or even entire cells. In simpler Vcell reaction diagrams, the inhibitor is typically not shown, since inhibition is built into the equation for the enzyme, represented by the node or yellow square in the figure above.
Let's now explore product inhibition in Vcell. In the reaction without product inhibition, R and Q are the reactant and product, respectively. S and P are used for the reaction with product P inhibition.
MODEL
Irreversible MM Kinetics - Without (left rx 1) and With (right, rx 2) Product Inhibition

Initial Conditions: No product inhibition
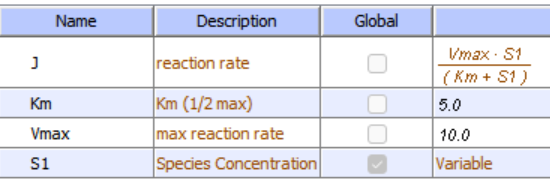
Initial Conditions: With product inhibition
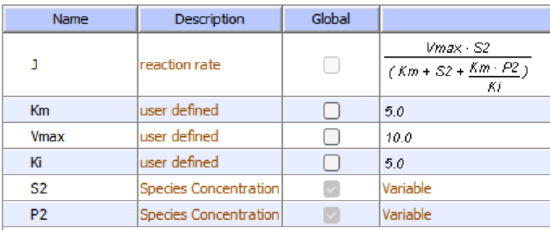
Select Load [model name] below
Select Start to begin the simulation.
Select Plot to change Y axis min/max, then Reset and Play | Select Slider to change which constants are displayed | Select About for software information.
Move the sliders to change the constants and see changes in the displayed graph in real-time.
Time course model made using Virtual Cell (Vcell), The Center for Cell Analysis & Modeling, at UConn Health. Funded by NIH/NIGMS (R24 GM137787); Web simulation software (miniSidewinder) from Bartholomew Jardine and Herbert M. Sauro, University of Washington. Funded by NIH/NIGMS (RO1-GM123032-04)
Inhibition by a competing substrate - the specificity constant
In the previous chapter, the specificity constant was defined as kcat/KM, which we also described as the second-order rate constant associated with the bimolecular reaction of E and S when S << KM. It also describes how well an enzyme distinguishes between substrates. If an enzyme encounters two different substrates, one can be considered a competitive inhibitor of the other. The following equation gives the ratio of initial velocities for two competing substrates at the same concentration equal to the ratio of their kcat/KM values.
\begin{equation}
\frac{\mathrm{v}_{\mathrm{A}}}{\mathrm{v}_{\mathrm{B}}}=\frac{\frac{\mathrm{k}_{\mathrm{catA}}}{\mathrm{K}_{\mathrm{A}}} \mathrm{A}}{\frac{\mathrm{k}_{\mathrm{cat} \mathrm{B}}}{\mathrm{K}_{\mathrm{B}}} \mathrm{B}}
\end{equation}
Here it is!
- Derivation
-
\begin{equation}
\mathrm{v}_{\mathrm{A}}=\frac{\mathrm{V}_{\mathrm{A}} \mathrm{A}}{\mathrm{K}_{\mathrm{A}}\left(1+\frac{\mathrm{B}}{\mathrm{K}_{\mathrm{B}}}\right)+\mathrm{A}} \quad \mathrm{v}_{\mathrm{B}}=\frac{\mathrm{V}_{\mathrm{B}} \mathrm{B}}{\mathrm{K}_{\mathrm{B}}\left(1+\frac{\mathrm{A}}{\mathrm{K}_{\mathrm{A}}}\right)+\mathrm{B}}
\end{equation}\begin{equation}
\frac{\mathrm{v}_{\mathrm{A}}}{\mathrm{v}_{\mathrm{B}}}=\frac{\frac{\mathrm{V}_{\mathrm{A}} \mathrm{A}}{\mathrm{K}_{\mathrm{A}}\left(1+\frac{\mathrm{B}}{\mathrm{K}_{\mathrm{B}}}\right)+\mathrm{A}}}{\frac{\mathrm{V}_{\mathrm{B}} \mathrm{B}}{\mathrm{K}_{\mathrm{B}}\left(1+\frac{\mathrm{A}}{\mathrm{K}_{\mathrm{A}}}\right)+\mathrm{B}}}=\frac{\frac{\mathrm{V}_{\mathrm{A}} \mathrm{A}}{\mathrm{K}_{\mathrm{A}}+\frac{\mathrm{K}_{\mathrm{A}} \mathrm{B}}{\mathrm{K}_{\mathrm{B}}}+\mathrm{A}}}{\frac{\mathrm{V}_{\mathrm{B}} \mathrm{B}}{\mathrm{K}_{\mathrm{B}}+\frac{\mathrm{K}_{\mathrm{B}} \mathrm{A}}{\mathrm{K}_{\mathrm{A}}}+\mathrm{B}}}
\end{equation}Now, in the above equation:
multiple the top half of the right-hand expression by \begin{equation}
\frac{\frac{1}{K_A}}{\frac{1}{K_A}}
\end{equation} multiple the bottom half of the right-hand expression by \begin{equation}
\frac{\frac{1}{K_B}}{\frac{1}{K_B}}
\end{equation} replace VA with kcatAE0 and VB with kcatBE0This gives the following expression for vA/vB:
\begin{equation}
\frac{\mathrm{v}_{\mathrm{A}}}{\mathrm{v}_{\mathrm{B}}}=\frac{\frac{\mathrm{k}_{\mathrm{catA}}}{\mathrm{K}_{\mathrm{A}}} \mathrm{A}}{\frac{\mathrm{k}_{\mathrm{cat} \mathrm{B}}}{\mathrm{K}_{\mathrm{B}}} \mathrm{B}}
\end{equation}
Uncompetitive Inhibition
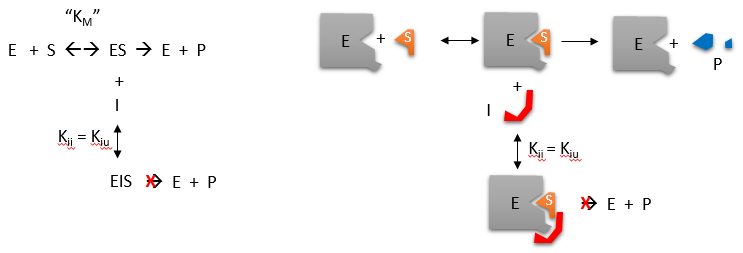
Reversible uncompetitive inhibition occurs when I binds only to ES and not free E. One can hypothesize that on binding S, a conformational change in E occurs, which presents a binding site for I. Inhibition occurs since ESI can not form the product. It is a dead-end complex that has only one fate: to return to ES. This is illustrated in the chemical equations and molecular cartoon shown in Figure \(\PageIndex{7}\).
\begin{equation}
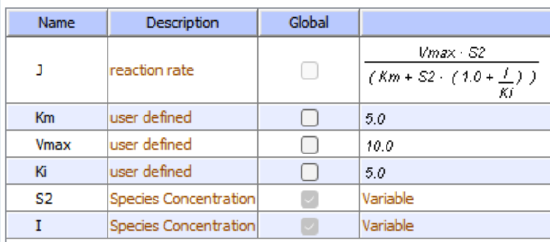
v_0=\frac{V_M S}{K_M+S\left(1+\frac{I}{K i i}\right)}=\frac{\left(\frac{V_M}{1+\frac{I}{K i i}}\right) S}{\left(\frac{K_M}{1+\frac{I}{K i i}}\right)+S}
\end{equation}
Let us assume that I binds reversibly to ES with a dissociation constant Kii. The second "i" in the subscript "ii" indicates that the intercept of the 1/v vs 1/S Lineweaver-Burk plot changes while the slope stays constant. Kii is also called Kiu, where the subscript "u" denotes the uncompetitive inhibition constant.
A look at the top mechanism shows that in the presence of I, as S increases to infinity, not all of E is converted to ES. There is a finite amount of ESI, even at an infinite S. Now remember that Vm = kcatE0 if and only if all E is in the form ES. Under these conditions, the apparent Vm, Vmapp, is less than the true Vm without the inhibitor. In addition, the apparent Km, Kmapp, will change. We can use LaChatelier's principle to understand this. If I binds to ES alone, and not E, it will shift the equilibrium of E + S ↔ ES to the right, which would have the effect of decreasing the Kmapp (i.e., it would appear that the affinity of E and S has increased). The double-reciprocal plot (Lineweaver-Burk plot) is a powerful way to visualize inhibition. In the presence of I, both Vm and Km decrease. Therefore, -1/Km, the x-intercept on the plot, will get more negative, and 1/Vm will get more positive. It turns out that they change to the same extent. Therefore, the plots will consist of a series of parallel lines, which is the hallmark of uncompetitive inhibition, as shown in Figure \(\PageIndex{8}\).

Here is an interactive graph showing v0 vs [S] for uncompetitive inhibition with Vm and Km both set to 100. Change the sliders for [I] and Kis and see the effect on the graph.
Here is an interactive graph showing uncompetitive inhibition with Vm and Km both set to 10. Change the sliders for [I] and Kii and see the effect on the graph
An equation shown in the diagram above can be derived to describe the effect of the uncompetitive inhibitor on the reaction velocity. The only change is that the S term in the denominator is multiplied by the factor 1+I/Kii. We want to rearrange this equation to show how Km and Vm are affected by the inhibitor, not S. Rearranging the equation above yields Kmapp = Km/(1 + I/Kii) and Vmapp = Vm/(1 + I/Kii). This shows that the apparent Km and Vm do decrease as we predicted. Kii is the inhibitor dissociation constant, in which the inhibitor affects the intercept of the double-reciprocal plot. Note that if I = 0, Km and Vm remain unchanged.
Progress Curves: Uncompetitive Inhibition
Now, let's compare the progress curves for an enzyme-catalyzed reaction in the absence and presence of an uncompetitive inhibitor.
MODEL
No inhibition (left) and Uncompetitive Inhibition (right)
(Note the the Vcell reaction diagram is the same as for competitive inhibition. However, the mathematical equations differ as shown below.
Initial values No Inhibition
Initial values With Uncompetitive Inhibitor
I is fixed for each simulation (as it is not converted to a product) but can be changed in the simulation below.
Select Load [model name] below
Select Start to begin the simulation.
Select Plot to change Y axis min/max, then Reset and Play | Select Slider to change which constants are displayed | Select About for software information.
Move the sliders to change the constants and see changes in the displayed graph in real-time.
Time course model made using Virtual Cell (Vcell), The Center for Cell Analysis & Modeling, at UConn Health. Funded by NIH/NIGMS (R24 GM137787); Web simulation software (miniSidewinder) from Bartholomew Jardine and Herbert M. Sauro, University of Washington. Funded by NIH/NIGMS (RO1-GM123032-04)
Figure \(\PageIndex{9}\) shows an interactive iCn3D model of an uncompetitive inhibitor of the cysteine protease caspase-6 (4HVA)
.png?revision=1&size=bestfit&width=353&height=323)
Figure \(\PageIndex{9}\): Uncompetitive inhibitor of the cysteine protease caspase-6 (4HVA) . (Copyright; author via source).
Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...TVfKio7NxcpX28
The "substrate" in this model (cyan, transparent surface, with labels on V, E, and I) is a substrate analog, VEI-CHO, in which the tripeptide substrate VEI ends not in a free carboxyl or amide group but in an aldehyde, which causes this "substrate" to become covalently attached to the enzyme and act as an inhibitor. The uncompetitive inhibitor (gray, transparent surface) binds to the blue surface. Hence, it binds to the ES complex. Two active site residues, Cys 163, the active site nucleophile, and His 121, a catalytic acid/base, are shown in colored sticks and labeled.
Figure \(\PageIndex{10}\) below shows a Lineweaver-Burk plot showing 1/vo vs 1/[S], where the substrate is the divalent compound (VEID)2R110. Its N-terminus is capped with a benzyloxy (Z) group. R110 is a rhodamine-type fluorophore, which, on cleavage, gives a strong fluorescent signal that the benzyloxy group initially quenches in the uncleaved substrate.
![A graph showing 1/velocity vs. 1/[(VEID)₂R110], with colored lines and dots representing various concentrations (0-100 nM).](https://bio.libretexts.org/@api/deki/files/76616/LBplotCasoace6_UncompInhib.svg?revision=1&size=bestfit&width=436&height=307)
Figure \(\PageIndex{10}\): Double-reciprocal Lineweaver-Burk plot of compound 3 with (VEID)2R110 substrate showing uncompetitive inhibition. Heise CE et al. (2012) Mechanistic and Structural Understanding of Uncompetitive Inhibitors of Caspase-6. PLoS ONE 7(12): e50864. https://doi.org/10.1371/journal.pone.0050864. Creative Commons Attribution License.
Noncompetitive and Mixed Inhibition
Reversible noncompetitive inhibition occurs when I binds to both E and ES. We will look at only the special case in which the dissociation constants of I for E and ES are the same. This is called noncompetitive inhibition. It is quite rare, as it would be difficult to imagine a large inhibitor that inhibits the turnover of a bound substrate and does not affect the binding of S to E. However, the covalent interaction of protons with both E and ES can lead to noncompetitive inhibition. In the more general case, the Kd's are different, and the inhibition is called mixed. Since inhibition occurs, we will hypothesize that ESI can not form the product. It is a dead-end complex with only one fate: to return to ES or EI. This is illustrated in the chemical equations and the molecular cartoon in Figure \(\PageIndex{11}\).
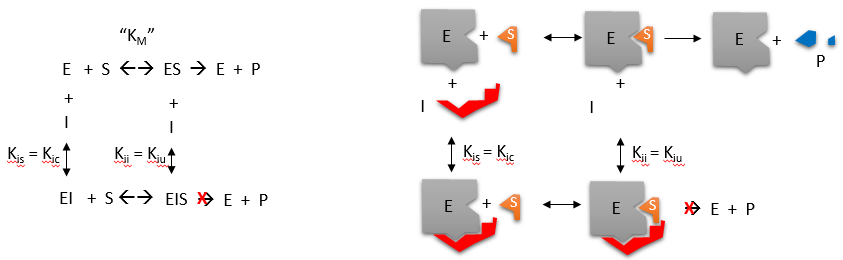
Let us assume for ease of equation derivation that I binds reversibly to E with a dissociation constant of Kis (as we denoted for competitive inhibition) and to ES with a dissociation constant Kii (as we noted for uncompetitive inhibition). Assume that for noncompetitive inhibition, Kis = Kii. A look at the top mechanism shows that in the presence of I, as S increases to infinity, not all of E is converted to ES. There is a finite amount of ESI, even at infinite S. Now remember that Vm = kcatE0 if and only if all E is in the form ES. Under these conditions, the apparent Vm, Vmapp, is less than the real Vm without an inhibitor. In contrast, the apparent Km, Kmapp, will not change since I binds to both E and ES with the same affinity and hence will not perturb that equilibrium, as deduced from LaChatelier's principle. The double-reciprocal plot (Lineweaver-Burk plot) is a powerful way to visualize inhibition. In the presence of I, just Vm will decrease. Therefore, -1/Km, the x-intercept will stay the same, and 1/Vm will become more positive. Therefore, the plots will consist of a series of lines intersecting on the x-axis, the hallmark of noncompetitive inhibition. You should be able to figure out how the plots would look if Kis differs from Kii (mixed inhibition).
\begin{equation}
v_0=\frac{V_M S}{K_M\left(1+\frac{I}{K i s}\right)+S\left(1+\frac{I}{K i i}\right)}
\end{equation}
The equation shown in the diagram above can be derived to describe the effect of a noncompetitive inhibitor on reaction velocity. Km is multiplied by 1+I/Kis, and S by 1+I/Kii in the denominator. We want to rearrange this equation to show how the inhibitor affects Km and Vm, not S. Rearranging the equation as shown above shows that Kmapp = Km(1+I/Kis)/(1+I/Kii) = Km when Kis = Kii, and Vmapp = Vm/(1+I/Kii). This shows that Km remains unchanged and that Vm decreases as we predicted. The plot shows a series of lines intersecting on the x-axis as shown in Figure \(\PageIndex{12}\). Both the slope and the y-intercept are changed, which are reflected in the names of the two dissociation constants, Kis and Kii. Note that if I is zero, Kmapp = Km and Vmapp = Vm. Sometimes, the Kis and Kii inhibition dissociation constants are called Kc and Ku (competitive and uncompetitive inhibition dissociation constants.
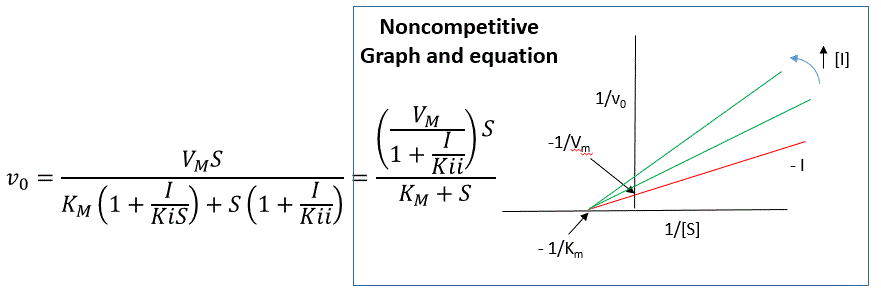
Move the sliders on this interactive graph below to show how changes in Kis and Kii affect the position on the graph where the lines intersect. Try changing their values to shift the graph intersections from the top-left quadrant to the bottom-left quadrant.
Here is an interactive graph showing v0 vs [S] for mixed inhibition with Vm and Km both set to 100. Change the sliders for [I] and Kis and see the effect on the graph
Here is an interactive graph showing mixed inhibition with Vm and Km both set to 10. Change the sliders for [I] and Kii and see the effect on the graph
Progress Curves: Mixed/Noncompetitive Inhibition
Now let's compare the progress curves for an enzyme-catalyzed reaction in the absence and presence of a mixed inhibitor.
MODEL
No inhibition (left) and Uncompetitive Inhibition (right)
Note that the Vcell reaction diagram is the same as for competitive and uncompetive inhibition. It doesn't explicitly show that the mixed inhibitor binds to both free and substrate-bound enzymes. Those interactions are addressed in the mathematical equations for mixed inhibition.
Initial values No Inhibition
Initial values With Uncompetitive Inhibitor
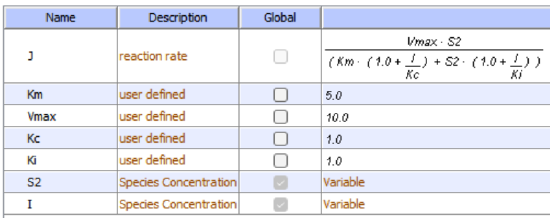
I is fixed for each simulation (as it is not converted to a product) but can be changed in the simulation below.
Select Load [model name] below
Select Start to begin the simulation.
Select Plot to change Y axis min/max, then Reset and Play | Select Slider to change which constants are displayed | Select About for software information.
Move the sliders to change the constants and see changes in the displayed graph in real-time.
Time course model made using Virtual Cell (Vcell), The Center for Cell Analysis & Modeling, at UConn Health. Funded by NIH/NIGMS (R24 GM137787); Web simulation software (miniSidewinder) from Bartholomew Jardine and Herbert M. Sauro, University of Washington. Funded by NIH/NIGMS (RO1-GM123032-04)
Mixed and noncompetitive inhibition (as shown by the mechanism above) differ from competitive and uncompetitive inhibition. Inhibitor binding is not simply a dead-end reaction in which the inhibitor can dissociate only in a single reverse step. In the above equilibrium, S can dissociate from ESI to form EI, so the system may not be at equilibrium. With dead-end steps, no flux of reactants occurs through the dead-end complex, so the equilibrium for the dead-end step is not perturbed.
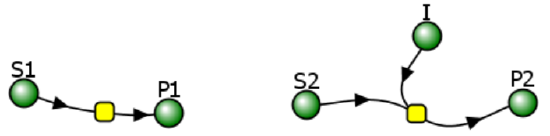
Other mechanisms can commonly give mixed inhibition. For example, the product released in a ping pong mechanism (discussed in the next chapter) can give mixed inhibition. A ping pong reaction mechanism is shown and superficially explained in Figure \(\PageIndex{13}\).
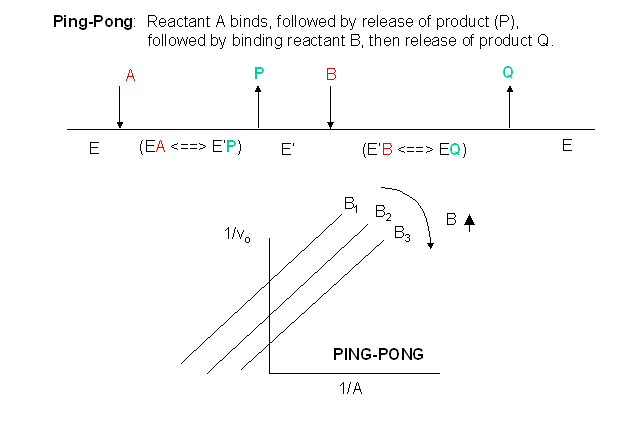
If P, acting as a product inhibitor, can bind to two different forms of the enzyme (E' and E), it will act as a mixed inhibitor.
Figure \(\PageIndex{14}\) shows an interactive iCn3D model of a noncompetitive inhibitor of M. tuberculosis's class IIa fructose 1,6-bisphosphate aldolase (4LV4).
.png?revision=1&size=bestfit&width=372&height=299)
Figure \(\PageIndex{14}\): Noncompetitive inhibitor of M. tuberculosis's class IIa fructose 1,6-bisphosphate aldolase (4LV4). (Copyright; author via source).
Click the image for a popup or use this external link: https://structure.ncbi.nlm.nih.gov/i...wZDtkvtwqnrVy7
The non-competitive inhibitor, 8-hydroxyquinoline carboxylic acid (HCA), is shown in cyan. Active-site side chains of the Zn2+-containing enzyme are shown as colored sticks, labeled.
The noncompetitive inhibitor does not enter into the pocket where the substrate (not shown in the model above) binds. The binding of the noncompetitive inhibitor causes a conformational change that moves the Zn-binding loop (Z-loop), which contains His 212, which coordinates the Zn2+ ion. Hence, the Zn-loop is part of the active site, and when it moves upon inhibitor binding, access to the empty pocket where the substrate binds is hindered. When Zn2+ moves away from the active site, it can no longer catalyze the reaction.
Enzyme Inhibition in Vivo
The pharmaceutical industry is devoted to finding drug molecules that affect biological processes. Typically, this means developing small-molecule inhibitors for target proteins. Recent work has expanded to include the development of inhibitory RNA molecules that affect DNA transcription and mRNA translation. Using combinatorial synthetic techniques and computational modeling, developing small-molecule inhibitors (especially competitive ones) that inhibit proteins in vitro using purified enzymes, substrates, and inhibitors in lab testing has become easier. Assuming that the inhibitor could pass through the membrane and accumulate to a sufficient concentration, would it have the same inhibitory properties in the cell as in the test tube? The answer turns out to be maybe. Remember that a cell is tightly packed with a multitude of other small molecules and macromolecules. In addition, the enzyme targeted for inhibition is most likely part of a pathway of enzymes that feed reactants into the enzyme and remove the product. Hence, the flux of substrate and product is controlled by the entire pathway and not just the single target enzyme. The target enzyme's product concentration is determined by the enzyme's kinetic parameters and the available substrate concentration.
The conditions under which enzymes are studied (in vitro) and those under which they operate (in vivo) are very different.
- In vitro (in the lab), the enzyme is held at a constant concentration while the substrate is varied (i.e., the substrate concentration is the independent variable). The velocity is determined by the substrate concentration. When inhibition is studied, the substrate is varied while the inhibitor is held constant at several different fixed concentrations.
- In vivo (in the cell), the velocity might be relatively fixed, with the substrate concentration determined by the velocity. The substrate can't build up at the enzyme to avoid a flux bottleneck, so the enzyme processes it in a steady-state manner to produce the product, as determined by the Michael-Menten equation.
What happens when an inhibitor is added in vivo? Let's assume that the enzyme is running at v = Vm/2. How might in vivo inhibition plots look at constant velocity (for example, v=Vm/2) when both I and S can vary, and in which S for an enzyme in the middle of a pathway is determined by v?
The equations and graph below show the ratio of S/Km vs I/Kix for inhibition at constant v, a condition encountered when an enzyme in a metabolic pathway is subject to flux controls imposed by the entire pathway. The x-axis reflects the ratio of inhibitor concentration to its inhibition constant. Likewise, the y-axis reflects the relative amount of substrate compared to its Km. The graph for in vivo competitive inhibition is linear, but it "blows up" for uncompetitive inhibition, as shown in Figure \(\PageIndex{15}\).

Here are the derivations used to produce the graphs in Figure \(\PageIndex{13}\).
Here it is!
- Derivation
-
Competitive Inhibition at constant velocity v:
Let's start with the equation of competitive inhibition.
\begin{equation}
\mathrm{v}=\frac{\mathrm{V}_{\mathrm{M}} \mathrm{S}}{\mathrm{K}_{\mathrm{M}}\left(1+\frac{\mathrm{I}}{\mathrm{K}_{\mathrm{is}}}\right)+\mathrm{S}}
\end{equation}Let
\begin{equation}
\left(1+\frac{\mathrm{I}}{\mathrm{K}_{\mathrm{is}}}\right)=\mathrm{y}
\end{equation}Then,
\begin{equation}
\begin{gathered}
v=\frac{V_M S}{K_M y+S} \\
v\left(K_M y+S\right)=V_M S \\
v K_M y+v S=V_M S \\
v K_M y=V_M S-v S=S\left(V_M-v\right) \\
K_M=\frac{S\left(V_M-v\right)}{v y}
\end{gathered}
\end{equation}which gives:
\begin{equation}
\begin{gathered}
\frac{S}{K_M}=\frac{v y}{\left(V_M-v\right)}=\frac{y}{\frac{V_M}{v}-1}=\frac{1+\frac{I}{K_{i s}}}{\frac{V_M}{v}-1}=\frac{1}{\frac{V_M}{v}-1}+\frac{\frac{I}{K_{i s}}}{\frac{V_M}{v}-1} \\
\frac{S}{K_M}=\frac{1}{\frac{V_M}{v}-1}+\left(\frac{1}{\frac{V_M}{v}-1}\right) \frac{I}{K_{i s}}
\end{gathered}
\end{equation}Note from the last equation that the graph of S/KM vs I/Kis is linear (at a fixed v), as shown in the above figure.
Uncompetitive Inhibition at constant velocity v:
Let's start with the equation of uncompetitive inhibition.
\begin{equation}
\mathrm{v}=\frac{\mathrm{V}_{\mathrm{M}} \mathrm{S}}{\mathrm{K}_{\mathrm{M}}+\mathrm{S}\left(1+\frac{\mathrm{I}}{\mathrm{K}_{\mathrm{ii}}}\right)}
\end{equation}Let
\begin{equation}
\left(1+\frac{1}{\mathrm{K}_{\mathrm{ii}}}\right)=\mathrm{y}
\end{equation}then:
\begin{equation}
\begin{gathered}
v=\frac{V_M S}{K_M+S y} \\
v\left(K_M+S y\right)=V_M S \\
v K_M=V_M S-v S y=S\left(V_M-v y\right)
\end{gathered}
\end{equation}which gives:
\begin{equation}
\frac{S}{K_M}=\frac{v}{V_M-v y}\left(\frac{\frac{1}{v}}{\frac{1}{v}}\right)=\frac{1}{\frac{V_M}{v}-y}=\frac{1}{v-1-\frac{I}{K_{i i}}}
\end{equation}This graph is not a linear function of I/Kii as it was for in vivo competitive inhibition.
- for competitive inhibition, the graph of S/KM is a linear function of I/Kis
- for uncompetitive inhibition, the graph of S/KM is NOT a linear function of I/Kii but rather "blows up" to infinity.
These graphs and associated equations are dramatically different from the very similar forms of inhibition equations and curves for in vitro inhibition at varying S and different fixed values of inhibitor. Consider the uncompetitive graph and equation. In the absence of an inhibitor, if S=Km, then Vm/v = 2, so the calculated value from the equation above of S/Km = 1. The y-intercept of the graph above is 1 for uncompetitive (and competitive) inhibition. If I is allowed to increase to a value of Kii (so I/Kii = 1), again at constant v=Vm/2, then the right-hand side goes to infinity.
In a linked series of reactions, if the middle reaction is inhibited, the substrate for that enzyme builds up, whether the inhibition is competitive or uncompetitive. In competitive inhibition, the substrate concentration can be increased to meet the enzyme's requirements. However, as the above figure shows, this can't happen with uncompetitive inhibition, since as more substrate accumulates, the reaction reaches a point at which steady state is lost.
Obviously, this limiting case can't be realistically reached, but it does suggest that uncompetitive inhibitors would be more effective in vivo in controlling a metabolic pathway than competitive inhibitors. Cornish-Bowden argues that purely uncompetitive inhibitors are rare because of the degree of inhibition they can hypothetically produce (1986). Likewise, he suggests that medicinal chemists should synthesize uncompetitive inhibitors if their goal is to maximally inhibit a metabolic pathway under the kind of flux control described above. Although it is more difficult to synthesize a purely uncompetitive inhibitor, as it can't be easily modeled after the structure of a natural ligand that binds to the active site, than competitive inhibitors, he notes that synthesizing mixed (and noncompetitive) inhibitors whose Kii values are of reasonable size compared to their Kis values would be one approach.
Move the sliders on the interactive graph below to show how the graphs change. Disregard the graph in the right lower quadrant, as that location would require negative values of either S or K.
Inhibition by Temperature and pH Changes
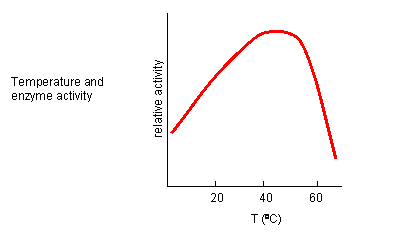
From 0 to about 40-50 oC, enzyme activity usually increases, as do the rates of most reactions in the absence of catalysts. (Remember the general rule of thumb that reaction velocities double for each increase of 10 oC). At higher temperatures, the enzyme's activity decreases dramatically as it denatures. These features are illustrated in Figure \(\PageIndex{16}\).
Likewise, pH has a marked effect on the velocity of enzyme-catalyzed reactions as illustrated in Figure \(\PageIndex{17}\).
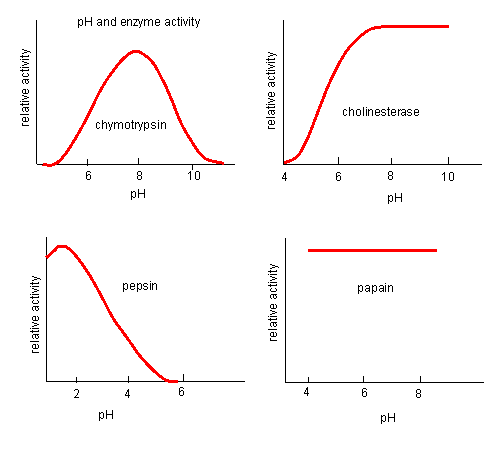
Think of all the things that pH changes might affect. It might
- affect E in ways to alter the binding of S to E, which would affect Km
- affect E in ways to alter the actual catalysis of bound S, which would affect kcat
- affect E by globally changing the conformation of the protein
- affect S by altering the protonation state of the substrate
The easiest assumption is that key side chains necessary for catalysis must be in the correct protonation state. Thus, a sidechain, with an apparent pKa of around 6, must be deprotonated for optimal trypsin activity, which shows a pH-dependent increase centered at pH 6. Which amino acid side chain would be a likely candidate?
Recent Updates: July 6, 2026
Figure \(\PageIndex{18}\) below shows linked chemical equations that could describe the effect of H+ on the kinetic parameters.

These equations assume that only the singly protonated form of the enzyme, EH and ESH, is active. At lower pH (more acidic), the doubly-protonated EH2+ and ESH2+ are inactive (perhaps due to the protonation of an essential catalytic base). Likewise, at higher pH (more basic), the fully deprotonated E- and ES- forms of the enzyme are inactive (perhaps due to deprotonation of an essential catalytic acid or a lack of a stabilizing positively charged, protonated species).
A series of equations shows how the kinetics (Vm app and Km app) depend on H+ (i.e., pH) and the pKas of the various protonated states of the enzyme.
For v0:
\begin{equation}
\mathrm{v}_{\mathrm{o}}=\frac{\mathrm{V}_{\mathrm{m} a p p} \mathrm{~S}}{\mathrm{~K}_{\mathrm{m} a p p}+\mathrm{S}}
\end{equation}
For the apparent Vm:
\begin{equation}
\mathrm{V}_{\mathrm{m} \text { app }}=\frac{\mathrm{V}_{\mathrm{m}}}{1+\frac{\mathrm{H}^{+}}{\mathrm{K}_{\mathrm{ES} 1}}+\frac{\mathrm{K}_{\mathrm{ES} 2}}{\mathrm{H}^{+}}}
\end{equation}
This can be rewritten as:
\begin{equation}
V_{m, a p p}=\frac{V_m}{1+10^{\left(p K a_1-p H\right)}+10^{\left(p H-p K a_2\right)}}
\end{equation}
Figure \(\PageIndex{19}\) below shows an interactive graph that allows you to change sliders to see how Vm app changes with pH and pKa1 and pka2. Note how the pKas can be deduced from the graphs.
p>
Figure \(\PageIndex{19}\): Interactive graph of Vm app vs pH with changing pKa1 and pka2 values. Graph made by Claude (Anthropic)
- Click for the derivation!
-
A diprotic enzyme–substrate complex in which only the singly protonated form is catalytically competent
Enzyme activity varies with pH because ionizable groups at the active site must be in a particular protonation state for catalysis to occur. The derivation below obtains the bell-shaped dependence of the apparent maximal velocity on pH for the simplest such case: a complex bearing two ionizable groups, one that must be deprotonated and one that must remain protonated.
1. The kinetic scheme
The enzyme and the enzyme–substrate complex each exist in three protonic states. Only the singly protonated species EH and ESH lie on the productive pathway:
E⁻ + S ⇌ ES⁻ (inactive)
EH + S ⇌ ESH → EH + P (active)
EH2⁺ + S ⇌ ESH2⁺ (inactive)
The horizontal steps of the middle row are governed by k1, k2 and k3; the vertical steps connecting the rows are the acid–base equilibria described by KE1, KE2 (free enzyme) and KES1, KES2 (enzyme–substrate complex).
Assumptions
(i) Rapid protonic equilibrium. Proton transfers are fast relative to catalysis, so every acid–base step remains at equilibrium throughout the reaction.
(ii) A single catalytically competent species. Only ESH proceeds to product; ES⁻ and ESH2⁺ are catalytically dead ends.
(iii) Irreversible product formation. The chemical step is irreversible with rate constant k3, and initial velocities are measured so that [P] ≈ 0.
2. Express the inactive complexes in terms of the active one
Write both ionizations of the ES complex as dissociations, each releasing a proton:
\[ K_{ES1}=\frac{[ESH][H^{+}]}{[ESH_{2}^{+}]} \qquad
K_{ES2}=\frac{[ES^{-}][H^{+}]}{[ESH]} \]Rearranging each expression to isolate the inactive species gives them in terms of [ESH] and the proton concentration:
\[ [ESH_{2}^{+}]=\frac{[ESH][H^{+}]}{K_{ES1}} \qquad
[ES^{-}]=\frac{K_{ES2}[ESH]}{[H^{+}]} \]Note: The two inactive forms respond to pH in opposite directions. [ESH2⁺] grows as the solution becomes more acidic ([H⁺] in the numerator), while [ES⁻] grows as it becomes more alkaline ([H⁺] in the denominator). This opposition is what ultimately produces a bell-shaped curve rather than a simple sigmoid.
3. Apply conservation of enzyme–substrate complex
The total concentration of complex in all three protonation states is the sum of the three species. Substituting the two results above and factoring out [ESH]:
\[ [ES]_{T}=[ESH]+[ESH_{2}^{+}]+[ES^{-}]
=[ESH]\left(1+\frac{[H^{+}]}{K_{ES1}}+\frac{K_{ES2}}{[H^{+}]}\right) \]The bracketed quantity recurs throughout the derivation, so it is convenient to name it the Michaelis pH function of the ES complex:
\[ f_{ES}=1+\frac{[H^{+}]}{K_{ES1}}+\frac{K_{ES2}}{[H^{+}]} \]
so that the concentration of the active complex is a simple fraction of the total:
\[ [ESH]=\frac{[ES]_{T}}{f_{ES}} \]
Interpretation: The quantity 1/fES is precisely the fraction of the enzyme–substrate complex present in the catalytically competent, singly protonated state. Because both added terms are positive, fES ≥ 1 always; it approaches 1 only at pH values where essentially all of the complex exists as ESH. Every subsequent result follows from this single idea.
4. Take the saturating-substrate limit
Because only ESH turns over, the observed velocity is
\[ v=k_{3}[ESH]=\frac{k_{3}[ES]_{T}}{f_{ES}} \]
At saturating substrate concentration, essentially all of the enzyme is driven into complex, so [ES]T → [E]T. This limiting velocity is by definition the apparent maximal velocity, and with Vm = k3[E]T we obtain the desired result:
\[ V_{m,app}=\frac{V_{m}}{1+\dfrac{[H^{+}]}{K_{ES1}}+\dfrac{K_{ES2}}{[H^{+}]}} \]
5. Why only the ES ionizations appear
A result worth emphasizing is what is absent from the equation. The free-enzyme ionization constants KE1 and KE2 do not appear at all. Saturating substrate removes free enzyme from the system, so the protonation state of E cannot influence the maximal velocity. Consequently:
• A V (or kcat) pH profile reports on ionizations in the enzyme–substrate complex.
• A V/K pH profile reports on ionizations in the free enzyme and free substrate.
• The Km pH profile depends on the ratio of both sets, and is flat only when substrate binding leaves the pKa values unperturbed.
This separation is the basis for using pH profiles to assign catalytic versus binding roles to active-site residues.
6. Conversion to pKₐ form
For plotting against pH it is convenient to eliminate [H⁺]. Substituting [H⁺] = 10−pH, KES1 = 10−pKa1 and KES2 = 10−pKa2 converts each ratio into a single power of ten:
\[ \frac{[H^{+}]}{K_{ES1}}=10^{(pK_{a1}-pH)} \qquad
\frac{K_{ES2}}{[H^{+}]}=10^{(pH-pK_{a2})} \]giving the form used for the interactive plot, in which pH is the independent variable and the two pKa values are parameters:
\[ V_{m,app}=\frac{V_{m}}{1+10^{(pK_{a1}-pH)}+10^{(pH-pK_{a2})}} \]
where pKa1 = −log KES1 and pKa2 = −log KES2. Note that −log[H⁺] is simply pH; the pKa values are parameters that position the two limbs, not the abscissa.
7. Position and height of the optimum
Because Vm,app is maximal where fES is minimal, differentiating fES with respect to pH and setting the derivative to zero gives 10(pKa1−pH) = 10(pH−pKa2). Equating exponents places the optimum at the midpoint of the two pKa values:
\[ pH_{opt}=\tfrac{1}{2}\left(pK_{a1}+pK_{a2}\right) \]
Substituting this pH back into the rate equation, and writing ΔpKa = pKa2 − pKa1 for the separation of the two groups, gives the peak height:
\[ V_{m,app}^{\,peak}=\frac{V_{m}}{1+2\cdot 10^{-\Delta pK_{a}/2}} \]
This makes quantitative an observation that is otherwise only qualitative: the observed maximum reaches the true Vm only when the two ionizations are well separated.
ΔpKa
peak Vm,app / Vm
width at half-maximum (pH units)
0
0.333
1.36
1
0.613
1.69
2
0.833
2.29
3
0.940
3.10
4
0.980
4.03
5
0.994
5.01
6
0.998
6.00
Once the two pKa values differ by about four units or more, the peak lies within 2% of Vm, and the half-maximal width converges on ΔpKa itself. Closely spaced ionizations both depress and narrow the curve, which is why pKa values fitted from closely spaced limbs require correction.
8. Limiting behavior of the two limbs
Far from the optimum, a single term dominates the denominator and the equation collapses to a simple exponential in pH. At low pH the term in KES1 dominates; at high pH the term in KES2 dominates:
\[ \text{low pH:}\;\; V_{m,app}\approx V_{m}\,10^{(pH-pK_{a1})} \qquad
\text{high pH:}\;\; V_{m,app}\approx V_{m}\,10^{(pK_{a2}-pH)} \]Taking logarithms shows that a plot of log Vm,app against pH has limiting slopes of +1 on the acidic limb and −1 on the basic limb. The intersection of these asymptotes with the log Vm plateau locates pKa1 and pKa2 graphically — the basis of the classical Dixon–Webb method of reading pKa values from a log-rate profile. Each limb is half-maximal at its corresponding pKa.
Now let's do the same for the apparent Km.
For the apparent Km:
\begin{equation}
K_{m, a p p}=K_m \cdot \frac{1+\frac{\left[H^{+}\right]}{K_{E 1}}+\frac{K_{E 2}}{\left[H^{+}\right]}}{1+\frac{\left[H^{+}\right]}{K_{E S 1}}+\frac{K_{E S 2}}{\left[H^{+}\right]}}
\end{equation}
This can be rewritten as:
\begin{equation}
K_{m, a p p}=K_m \cdot \frac{1+10^{\left(p K a_{E 1}-p H\right)}+10^{\left(p H-p K a_{E 2}\right)}}{1+10^{\left(p K a_{E S 1}-p H\right)}+10^{\left(p H-p K a_{E S 2}\right)}}
\end{equation}
Figure \(\PageIndex{20}\) below shows an interactive graph that allows you to change sliders to see how Km app changes with pH and pKaE1, pKaE2, pKaES1, pKaES2 values.
p>
Figure \(\PageIndex{20}\): Interactive graph of Km app vs pH with changing pKaE1, pKaE2, pKaES1, pKaES2 values. Graph made by Claude (Anthropic)
- Click here for the derivation
-
How ionizations of the free enzyme and of the enzyme–substrate complex combine to determine Km,app
The apparent maximal velocity depends only on ionizations of the ES complex, because saturating substrate removes free enzyme from the system. The Michaelis constant is different: it is a ratio of bound to free states, so both sets of ionization constants survive. The derivation below obtains that result, identifies the three plateaus it predicts, and shows the special case in which the pH dependence disappears entirely.
1. The kinetic scheme
Both the free enzyme and the enzyme–substrate complex exist in three protonic states. Only the singly protonated species are productive: EH is the only form that binds substrate, and ESH is the only form that turns over.
E⁻ + S ⇌ ES⁻ (inactive)
EH + S ⇌ ESH → EH + P (active)
EH2⁺ + S ⇌ ESH2⁺ (inactive)
The left-hand column is connected by the free-enzyme ionization constants KE1 and KE2; the right-hand column by the complex ionization constants KES1 and KES2. These two sets are colored throughout this document to keep them distinct.
Assumptions
(i) Rapid equilibrium. Substrate binding and all proton transfers are fast relative to catalysis, so every binding and acid–base step remains at equilibrium. The intrinsic Michaelis constant is then the true dissociation constant, Km = [EH][S]/[ESH].
(ii) A single binding-competent and a single catalytically competent species. Only EH binds S, and only ESH proceeds to product.
(iii) Irreversible product formation, with rate constant k3, and initial velocities measured so that [P] ≈ 0.
2. Collect each column into a single pH function
Each ionization is written as a dissociation releasing a proton, so that every minor species can be expressed in terms of its singly protonated parent:
\[ [EH_{2}^{+}]=\frac{[EH][H^{+}]}{K_{E1}} \qquad
[E^{-}]=\frac{K_{E2}[EH]}{[H^{+}]} \]and identically for the complex:
\[ [ESH_{2}^{+}]=\frac{[ESH][H^{+}]}{K_{ES1}} \qquad
[ES^{-}]=\frac{K_{ES2}[ESH]}{[H^{+}]} \]Summing each column gives the total free enzyme and the total complex, each as its active form multiplied by a Michaelis pH function:
\[ \sum[E]=[EH]\,f_{E} \qquad \sum[ES]=[ESH]\,f_{ES} \]
\[ f_{E}=1+\frac{[H^{+}]}{K_{E1}}+\frac{K_{E2}}{[H^{+}]} \qquad
f_{ES}=1+\frac{[H^{+}]}{K_{ES1}}+\frac{K_{ES2}}{[H^{+}]} \]Interpretation 1/fE and 1/fES are the fractions of free enzyme and of complex that are in the singly protonated state. Both are ≤ 1, and each equals 1 only where its column is essentially all in the active protonation state. The entire pH behavior of the kinetic parameters follows from these two fractions.
3. Conservation and the rate equation
Total enzyme is distributed over both columns:
\[ [E]_{T}=[EH]\,f_{E}+[ESH]\,f_{ES} \]
The rapid-equilibrium binding step supplies [EH] = Km[ESH]/[S]. Substituting and factoring out [ESH]:
\[ [E]_{T}=[ESH]\left(\frac{K_{m}f_{E}}{[S]}+f_{ES}\right) \]
Since only ESH turns over, v = k3[ESH]. Solving for [ESH] and multiplying numerator and denominator by [S] gives
\[ v=\frac{k_{3}[E]_{T}[S]}{K_{m}f_{E}+f_{ES}[S]} \]
4. Recover the Michaelis–Menten form
Dividing numerator and denominator by fES puts the rate law back into standard hyperbolic form:
\[ v=\frac{\dfrac{k_{3}[E]_{T}}{f_{ES}}\,[S]}{\dfrac{K_{m}f_{E}}{f_{ES}}+[S]} \]
Comparing term by term with v = Vm,app[S] / (Km,app + [S]) identifies both apparent parameters at once:
\[ V_{m,app}=\frac{k_{3}[E]_{T}}{f_{ES}}=\frac{V_{m}}{f_{ES}} \qquad
K_{m,app}=K_{m}\,\frac{f_{E}}{f_{ES}} \]Written out in full, the apparent Michaelis constant is therefore
\[ K_{m,app}=K_{m}\,
\frac{1+\dfrac{[H^{+}]}{K_{E1}}+\dfrac{K_{E2}}{[H^{+}]}}
{1+\dfrac{[H^{+}]}{K_{ES1}}+\dfrac{K_{ES2}}{[H^{+}]}} \]The key structural point: the numerator carries the free-enzyme constants and the denominator carries the complex constants. This asymmetry is not arbitrary: Km is a ratio of free to bound enzyme, so ionizations that deplete the free enzyme raise it while ionizations that deplete the complex lower it. Contrast this with Vm,app = Vm/fES, which contains no free-enzyme constants at all.
5. The special case in which the pH dependence vanishes
If substrate binding does not perturb the ionizable groups, then the complex ionizes exactly as the free enzyme does:
\begin{equation}
K_{E 1}=K_{E S 1}, \quad K_{E 2}=K_{E S 2} \Rightarrow f_E=f_{E S}
\end{equation}The two trinomials are then identical; they cancel, and the apparent Michaelis constant reduces to a pH-independent constant:
\[ K_{m,app}=K_{m}\,\frac{f_{E}}{f_{ES}}=K_{m} \qquad \text{(when } K_{E1}=K_{ES1},\; K_{E2}=K_{ES2}\text{)} \]
Worth flagging to students: A flat Km versus pH profile is therefore not a trivial or uninformative result — it is a positive statement that substrate binding leaves the pKa values of the ionizing groups unchanged. Any curvature in the profile is a direct measure of how far binding shifts them. Note also that an equation written with the same pair of constants in numerator and denominator is algebraically just Km, and will plot as a horizontal line.
6. Conversion to pKₐ form
Substituting [H⁺] = 10−pH and K = 10−pKa for each of the four constants converts every ratio into a single power of ten, giving the form used for plotting against pH:
\[ K_{m,app}=K_{m}\,
\frac{1+10^{(pK_{a,E1}-pH)}+10^{(pH-pK_{a,E2})}}
{1+10^{(pK_{a,ES1}-pH)}+10^{(pH-pK_{a,ES2})}} \]where pKa,E1 = −log KE1 and so on. As before, −log[H⁺] is simply pH; the four pKa values are parameters that position the transitions.
7. The three plateaus
Unlike the bell-shaped Vm,app profile, this expression is a ratio of two similar functions, so it does not peak — it steps between plateaus. At each pH extreme a single term dominates both numerator and denominator, and the ratio collapses to a constant.
At low pH the acidic terms dominate top and bottom, and the powers of ten combine:
\[ K_{m,app}\approx K_{m}\,\frac{10^{(pK_{a,E1}-pH)}}{10^{(pK_{a,ES1}-pH)}}
=K_{m}\,10^{(pK_{a,E1}-pK_{a,ES1})} \]and by the same argument at high pH:
\[ K_{m,app}\approx K_{m}\,10^{(pK_{a,ES2}-pK_{a,E2})} \]
In the intermediate range, both trinomials approach unity and Km,app → Km. The pH dependence therefore reduces to three readable levels:
pH regime
limiting Km,app
meaning
example value
low pH
Km · 10(pKa,E1 − pKa,ES1)
shift of the acidic group on binding
500
intermediate
Km
intrinsic binding constant
50
high pH
Km · 10(pKa,ES2 − pKa,E2)
shift of the basic group on binding
500
Example values are for Km = 50, pKa,E = 5 and 8, pKa,ES = 4 and 9. Each plateau is a direct read-out of a pKa shift on binding: one log unit of shift is one decade of change in Km,app. A plateau may lie above or below Km depending on the direction of the shift.
Because each plateau ratio is fixed by a difference of pKa values, it is also a statement of the thermodynamic cycle linking protonation and binding: the free energy of substrate binding and the free energies of ionization of E and ES are not independent, and the plateau height measures exactly that coupling.
The specificity constant, Vm/Km, as a function of pH and pKas.
The constant kcat/KM is also called the specificity constant, as it describes how well an enzyme distinguishes between competing substrates. We discussed this in the previous chapter section. How is it altered by pH? That is, how are Vm app/Km app affected by pH and pKas?
Dividing the two apparent parameters obtained in Section 4 gives a result that is less complicated than either one alone:
\[ \frac{V_{m,app}}{K_{m,app}}
=\frac{V_{m}/f_{ES}}{K_{m}\,f_{E}/f_{ES}}
=\frac{V_{m}}{K_{m}\,f_{E}} \]
The complex pH function cancels completely. The specificity constant therefore depends on only the free-enzyme ionizations, and traces a simple bell-shaped curve in fE — the same functional form as the Vm,app profile, but governed by the other set of constants:
\[ \left(\frac{V_{m}}{K_{m}}\right)^{app}
=\frac{V_{m}/K_{m}}{1+10^{(pK_{a,E1}-pH)}+10^{(pH-pK_{a,E2})}} \]
The three profiles together:
- A V profile reports ionizations of the ES complex;
- a V/K profile reports ionizations of the free enzyme; and
- a Km profile reports the difference between the two, which is why it is the only one of the three that can be flat.
Figure \(\PageIndex{21}\) below shows an interactive graph that allows you to change sliders to see how Vm app/Km app, (the apparent specificity constant) changes with pH and pKa1 and pka2.
p>
Figure \(\PageIndex{21}\): An interactive graph that allows you to change sliders to see how Vm app/Km app, (the apparent specificity constant) changes with pH and pKa1 and pka2. Graph made by Claude (Anthropic)
Summary
(Summary written by Claude, Sonnet 4.6, Anthropic)
This chapter develops the quantitative framework for enzyme inhibition — the reduction of catalytic activity by molecules that bind to the enzyme — covering the major reversible inhibition mechanisms, their mathematical descriptions, their Lineweaver-Burk signatures, special cases including product inhibition and substrate competition, and the physiologically important differences between inhibition analyzed in vitro versus in vivo.
Irreversible covalent inhibition provides a conceptual baseline: reagents that chemically modify essential active site residues (for example, iodoacetamide, which alkylates a catalytic Cys) permanently inactivate the enzyme. Distinguishing which residue is critical requires protection experiments in which a saturating ligand occupies the active site during chemical modification, with the extent of protection depending on KD and ligand concentration. However, the majority of pharmacologically relevant inhibition is reversible and noncovalent.
Competitive inhibition occurs when inhibitor I and substrate S compete for the same binding site on free enzyme E, binding in a mutually exclusive fashion (either through direct overlap of binding sites or through allosteric conformational changes that make one site inaccessible when the other is occupied). The key kinetic consequences follow from Le Chatelier's principle: I binding to free E shifts the E + S ↔ ES equilibrium leftward, increasing the apparent KM. However, at infinite [S], all E is driven into the ES form regardless of I concentration, so VM is unchanged. The modified velocity equation v₀ = VM[S]/(KM(1 + I/KIS) + [S]) shows that KMapp = KM(1 + I/KIS) grows with increasing I while VM is unaffected. On Lineweaver-Burk plots, competitive inhibition produces a family of lines with the same y-intercept (1/VM) but progressively more positive slopes (KM(1 + I/KIS)/VM) and x-intercepts moving toward zero (−1/KMapp). Competitive inhibitors are typically substrate analogs that mimic the structure of S, as illustrated by the buffer component MES, which acts as a competitive inhibitor of phosphotyrosyl phosphatase by mimicking the phosphate group of phosphotyrosine substrates.
Uncompetitive inhibition occurs when I binds only to the ES complex, not to free E — perhaps because substrate binding induces a conformational change that creates the inhibitor binding site. Since I cannot bind free E, it cannot compete with S for E; instead, it traps ES in a dead-end ESI complex. By Le Chatelier's principle, I binding to ES shifts the E + S ↔ ES equilibrium rightward, which paradoxically decreases KMapp = KM/(1 + I/Kii) while also decreasing VMapp = VM/(1 + I/Kii) — both change by the same factor (1 + I/Kii). On Lineweaver-Burk plots, this produces the hallmark of uncompetitive inhibition: a family of parallel lines (constant slope KM/V,, but shifted intercepts), distinguishing it from competitive inhibition where lines converge on the y-axis.
Noncompetitive inhibition is a special case of mixed inhibition in which I binds to both E and ES with equal dissociation constants KIS = Kii. Since I binds both forms equally, it does not perturb the E + S ↔ ES equilibrium, and KM is unchanged. However, since ESI is a dead-end complex that cannot produce product, VMapp = VM/(1 + I/Kii) is reduced. On Lineweaver-Burk plots, noncompetitive inhibition gives lines with a common x-intercept (−1/KM, since KM is unchanged) but increasing y-intercepts (1/VMapp), distinguishing it from competitive and uncompetitive cases. In mixed inhibition (the more general case where KIS ≠ Kii), both KMapp = KM(1 + I/KIS)/(1 + I/Kii) and VMapp = VM/(1 + I/Kii) change, and Lineweaver-Burk lines intersect in a quadrant displaced from either axis. The specificity constant kcat/KM governs competition between substrates: when two substrates compete for the same active site, the ratio of their initial velocities at equal concentration equals the ratio of their kcat/KM values, explaining how an enzyme discriminates among substrates in vivo.
Product inhibition — a special case of competitive inhibition in which the structurally similar product P occupies the active site — occurs in most enzymes to some degree and causes downward curvature in P vs. t progress curves as product accumulates. Tight product binding can significantly underestimate measured v₀ if initial rates are not determined carefully. The ping-pong reaction mechanism (two-substrate, two-product reactions with a covalent intermediate) can also generate mixed inhibition when the released product can bind to two different enzyme forms.
The physiological significance of inhibitor type is dramatically revealed by in vivo analysis. In the test tube, [S] is the independent variable and [I] is held constant. In the cell, however, an enzyme embedded in a metabolic pathway operates under approximately constant flux v, and [S] adjusts to satisfy the Michaelis-Menten equation rather than being experimentally controlled. Under constant flux conditions, deriving S/KM as a function of I/KIS (competitive) or I/Kii (uncompetitive) reveals a striking difference: competitive inhibition gives a linear S/KM vs. I/KIS relationship (the substrate concentration rises proportionally to compensate for the inhibitor), while uncompetitive inhibition makes S/KM go to infinity as I → Kii — meaning that no amount of substrate buildup can restore the required flux. This analysis, originally articulated by Cornish-Bowden, implies that uncompetitive inhibitors (and mixed inhibitors with relatively small Kii compared to KIS) would be far more effective at disrupting metabolic pathways in vivo than competitive inhibitors — a counterintuitive conclusion with important implications for drug design, since competitive inhibitors are most commonly developed due to their structural similarity to natural substrates.
Finally, pH affects enzyme activity by altering the protonation state of catalytically essential residues — typically requiring specific protonation states for general acid/base catalysis (affecting kcat) or for electrostatic interactions that position the substrate (affecting KM) — and produces characteristic bell-shaped activity-pH profiles whose inflection points reflect the apparent pKa values of critical active site residues. Temperature increases enzyme activity up to ~40–50°C via standard Arrhenius kinetics but causes a precipitous drop in activity above this range as the protein denatures, reflecting the thermodynamic marginal stability of folded proteins discussed in earlier chapters.