
4.8: Photosynthesis - The Role of Light
- Page ID
- 4634

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)The heart of photosynthesis as it occurs in most autotrophs consists of two key processes:
- the removal of hydrogen (H) atoms from water molecules
- the reduction of carbon dioxide (CO2) by these hydrogen atoms to form organic molecules.
The second process involves a cyclic series of reactions named (after its discoverer) the Calvin Cycle.
The electrons (e−) and protons (H+) that make up hydrogen atoms are stripped away separately from water molecules.
\[\ce{2H2O -> 4e^{-} + 4H^{+} + O2}\]
The electrons serve two functions:
- They reduce NADP+ to NADPH for use in the Calvin Cycle.
- They set up an electrochemical charge that provides the energy for pumping protons from the stroma of the chloroplast into the interior of the thylakoid.
The protons also serve two functions:
- They participate in the reduction of NADP+ to NADPH.
- As they flow back out from the interior of the thylakoid (by facilitated diffusion), passing down their concentration gradient), the energy they give up is harnessed to the conversion of ADP to ATP.
- Because it is drive by light, this process is called photophosphorylation.
\[\ce{ADP + P_i -> ATP}\]
The ATP provides the second essential ingredient for running the Calvin Cycle.
The removal of electrons from water molecules and their transfer to NADP+ requires energy. The electrons are moving from a redox potential of about +0.82 volt in water to −0.32 volt in NADPH. Thus enough energy must be available to move them against a total potential of 1.14 volts. Where does the needed energy come from? The answer: Light.
The Thylakoid Membrane
Chloroplasts contain a system of thylakoid membranes surrounded by a fluid stroma. Six different complexes of integral membrane proteins are embedded in the thylakoid membrane. The exact structure of these complexes differs from group to group (e.g., plant vs. alga) and even within a group (e.g., illuminated in air or underwater). They are as follows:
Photosystem I
The structure of photosystem I in a cyanobacterium ("blue-green alga") has been completely worked out. It probably closely resembles that of plants as well. It is a homotrimer with each subunit in the trimer containing:
- 12 different protein molecules bound to
- 96 molecules of chlorophyll a
- 2 molecules of the reaction center chlorophyll P700
- 4 accessory molecules closely associated with them
- 90 molecules that serve as antenna pigments
- 22 carotenoid molecules
- 4 lipid molecules
- 3 clusters of Fe4S4
- 2 phylloquinones
Photosystem II
Photosystem II is also a complex of
- > 20 different protein molecules bound to
- 50 or more chlorophyll a molecules
- 2 molecules of the reaction center chlorophyll P680
- 2 accessory molecules close to them
- 2 molecules of pheophytin (chlorophyll without the Mg++)
- the remaining molecules of chlorophyll a serve as antenna pigments.
- some half dozen carotenoid molecules. These also serve as antenna pigments.
- 2 molecules of plastoquinone
Light-Harvesting Complexes (LHC)
- LHC-I associated with photosystem I
- LHC-II associated with photosystem II
These LHCs also act as antenna pigments harvesting light and passing its energy on to their respective photosystems.
The LHC-II of spinach is a homotrimer, with each monomer containing
- a single polypeptide
- 8 molecules of chlorophyll a
- 6 molecules of chlorophyll b
- 4 carotenoid molecules
Cytochromes b6 and f
ATP synthase
How the System Works
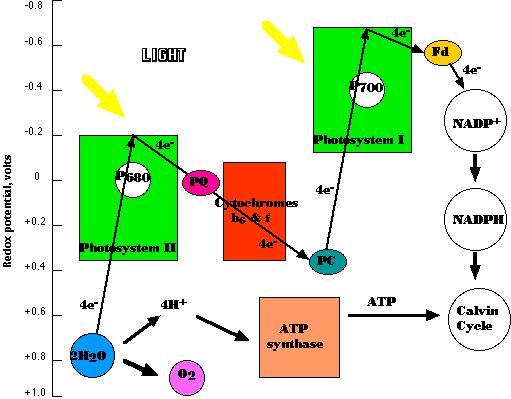
- Light is absorbed by the antenna pigments of photosystems II and I.
- The absorbed energy is transferred to the reaction center chlorophylls, P680 in photosystem II, P700 in photosystem I.
- Absorption of 1 photon of light by Photosystem II removes 1 electron from P680.
- With its resulting positive charge, P680 is sufficiently electronegative that it can remove 1 electron from a molecule of water.
- When these steps have occurred 4 times, requiring 2 molecules of water, 1 molecule of oxygen and 4 protons (H+) are released
- The electrons are transferred (by way of plastoquinone — PQ in the figure) to the cytochrome b6/f complex where they provide the energy for chemiosmosis.
- Activation of P700 in photosystem I enables it to pick up electrons from the cytochrome b6/f complex (by way of plastocyanin — PC in the figure) and raise them to a sufficiently high redox potential that, after passing through ferredoxin (Fd in the figure),
- they can reduce NADP+ to NADPH.
The sawtooth shifts in redox potential as electrons pass from P680 to NADP+ have caused this system to be called the Z-Scheme (although as I have drawn the diagram, it looks more like an "N"). It is also called noncyclic photophosphorylation because it produces ATP in a one-way process (unlike cyclic photophosphorylation and pseudocyclic photophosphorylation described below).
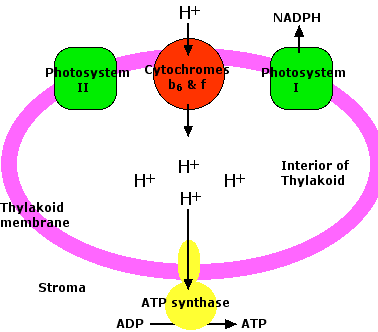
Chemiosmosis in Chloroplasts
The energy released as electrons pass down the gradient between photosystem II and plastocyanin (PC) is harnessed by the cytochrome b6/f complex to pump protons (H+) against their concentration gradient from the stroma of the chloroplast into the interior of the thylakoid (an example of active transport). As their concentration increases inside (which is the same as saying that the pH of the interior decreases), a strong diffusion gradient is set up. The only exit for these protons is through the ATP synthase complex. As in mitochondria, the energy released as these protons flow down their gradient is harnessed to the synthesis of ATP. The process is called chemiosmosis and is an example of facilitated diffusion.
-rev3.png?revision=1)
Cyclic Photophosphorylation
- Each CO2 taken up by the Calvin cycle) requires 2 NADPH molecules and 3 ATP molecules
- Each molecule of oxygen released by the light reactions supplies the 4 electrons needed to make 2 NADPH molecules.
- The chemiosmosis driven by these 4 electrons as they pass through the cytochrome b6/f complex liberates only enough energy to pump 12 protons into the interior of the thylakoid.
- But in order to make 3 molecules of ATP, the ATPase in chloroplasts appears to have 14 protons (H+) pass through it.
- So there appears to be a deficit of 2 protons.
- How is this deficit to be made up?
- One likely answer: cyclic photophosphorylation.
In cyclic photophosphorylation,
- the electrons expelled by the energy of light absorbed by photosystem I pass, as normal, to ferredoxin (Fd).
- But instead of going on to make NADPH,
- they pass to plastoquinone (PQ) and on back into the cytochrome b6/f complex.
- Here the energy each electron liberates pumps 2 protons (H+) into the interior of the
- thylakoid — enough to make up the deficit left by noncyclic photophosphorylation.
This process is truly cyclic because no outside source of electrons is required. Like the photocell in a light meter, photosystem I is simply using light to create a flow of current. The only difference is that instead of using the current to move the needle on a light meter, the chloroplast uses the current to help synthesize ATP.
Pseudocyclic Photophosphorylation
Another way to make up the deficit is by a process called pseudocyclic photophosphorylation in which some of the electrons passing to ferredoxin then reduce molecular oxygen back to H2O instead of reducing NADP+ to NADPH.
At first glance, this might seem a fruitless undoing of all the hard work of photosynthesis. But look again. Although the electrons cycle from water to ferredoxin and back again, part of their pathway is through the chemiosmosis-generating stem of cytochrome b6/f. Here, then, is another way that simply by turning on a light, enough energy is imparted to electrons that they can bring about the synthesis of ATP.
Antenna Pigments
Chlorophylls a and b differ slightly in the wavelengths of light that they absorb best (although both absorb red and blue much better than yellow and green). Carotenoids help fill in the gap by strongly absorbing green light. The entire complex ensures that most of the energy of light will be trapped and passed on to the reaction center chlorophylls.