
19.3B: Influenza
- Page ID
- 5978

Influenza is a viral infection of the lungs characterized by fever, cough, and severe muscle aches. In the elderly and infirm, it is a major cause of disability and death (often as a result of secondary infection of the lungs by bacteria). Even in the young and healthy, influenza produces a prostrating disease of a few days duration and one not soon forgotten. Influenza is not a case of low fever and sniffles that keeps you home in bed for a day nor a gastrointestinal upset ("stomach flu").
Influenza was responsible for the most devastating plague in human history - the "Spanish" flu that swept around the world in 1918 killing 675,000 people in the U.S. and an estimated 20–50 million people worldwide. (A disease that attacks a large fraction of the population in every region of the world is called a pandemic.) (It is uncertain where the flu first appeared, but it certainly wasn't in Spain.)
No one at the time even knew what disease agent was causing the pandemic. Not until 1930 (in pigs) and 1933 (in humans) was it established that influenza is caused by a virus.
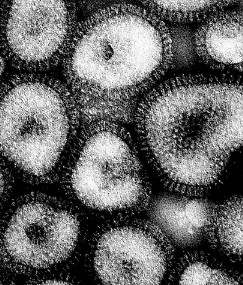
This electron micrograph (courtesy of Dr. K. G. Murti) shows several influenza virus particles (at a magnification of about 265,000x). The surface projections are molecules of hemagglutinin and neuraminidase (see below).
There are three types of influenza:
- Common but seldom causes disease symptoms.
- Often causes sporadic outbreaks of illness, especially in residential communities like nursing homes.
- Responsible for regular outbreaks, including the one of 1918. Influenza A viruses also infect domestic animals (pigs, horses, chickens, ducks) and some wild birds.
The Influenza A Virus
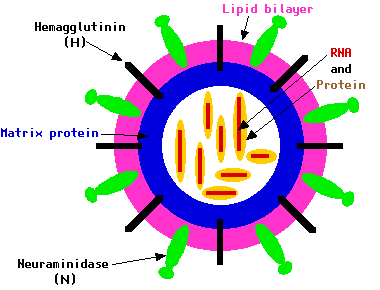
The influenza A virion is
- a globular particle (about 100 nm in diameter)
- sheathed in a lipid bilayer (derived from the plasma membrane of its host)
- Studded in the lipid bilayer are three integral membrane proteins
- some 500 molecules of hemagglutinin ("H")
- some 100 molecules of neuraminidase ("N")
- the M2 membrane protein (not shown).
- Encased by the lipid bilayer are
- some 3000 molecules of matrix protein
- 8 pieces or segments of RNA
Each of the 8 RNA molecules is associated with
- many copies of a nucleoprotein
- the three subunits of its RNA polymerase
- some "non-structural" protein molecules of uncertain function
The Genes of Influenza A
The 8 RNA molecules (the number in brackets is the designated segment number):
- The HA gene [4] encodes the hemagglutinin. 3 distinct hemagglutinins (H1, H2, and H3) are found in human infections; 15 others have been found in animal flu viruses.
- The NA gene [6] encodes the neuraminidase. 2 different neuraminidases (N1 and N2) have been found in human viruses; 9 others in other animals.
- The NP gene [5] encodes the nucleoprotein. Influenza A, B, and C viruses have different nucleoproteins.
- The M gene [7] encodes two proteins (using different reading frames of the RNA): a matrix protein (M1 — shown in blue) and an ion channel (M2) spanning the lipid bilayer (not shown).
- The NS gene [8] encodes two different non-structural proteins (also by using different reading frames). These are found in the cytosol of the infected cell but not within the virion itself.
- One RNA molecule (PA [3], PB1 [2], PB2 [1]) encoding each of the 3 subunits of the RNA polymerase. Occasional frameshifts during translation of PA produce a protein that reduces host gene expression.
The Disease
The influenza virus invades cells of the respiratory passages.
- Its hemagglutinin molecules bind to sialic acid residues on the glycoproteins exposed at the surface of the epithelial cells of the host respiratory system.
- The virus is engulfed by receptor mediated endocytosis.
- The drop in pH in the endosome (endocytic vesicle) produces a change in the structure of the viral hemagglutinin enabling it to
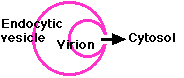
- fuse the viral membrane with the vesicle membrane.
- This exposes the contents of the virus to the cytosol.
- The RNA enter the nucleus of the cell where fresh copies are made.
- These return to the cytosol where some serve as messenger RNA (mRNA) molecules to be translated into the proteins of fresh virus particles.
- Fresh virus buds off from the plasma membrane of the cell (aided by the M2 protein) thus
- spreading the infection to new cells.
The result is a viral pneumonia. It usually does not kill the patient (the 1918 pandemic was an exception; some victims died within hours) but does expose the lungs to infection by various bacterial invaders that can be lethal. Before the discovery of the flu virus, the bacterium Hemophilus influenzae was so often associated with the disease that it gave it its name.
Pandemics and Antigenic Shift
Three pandemics of influenza have swept the world since the "Spanish" flu of 1918.
- The "Asian" flu pandemic of 1957;
- the "Hong Kong" flu pandemic of 1968;
- the "Swine" flu pandemic that began in April of 2009.
The pandemic of 1957 probably made more people sick than the one of 1918. But the availability of antibiotics to treat the secondary infections that are the usual cause of death resulted in a much lower death rate. The hemagglutinin of the 1918 flu virus was H1, its neuraminidase was N1, so it is designated as an H1N1 "subtype". Here are some others.
| Date | Strain | Subtype | Notes |
|---|---|---|---|
| 1918 | H1N1 | pandemic of "Spanish" flu | |
| 1957 | A/Singapore/57 | H2N2 | pandemic of "Asian" flu |
| 1962 | A/Japan/62 | H2N2 | epidemic |
| 1964 | A/Taiwan/64 | H2N2 | epidemic |
| 1968 | A/Aichi/68 | H3N2 | pandemic of "Hong Kong" flu |
| 1976 | A/New Jersey/76 | H1N1 | swine flu in recruits |
| 1977 | A/USSR/77 | H1N1 | "Russian" flu |
| 2009 | A/California/09 | H1N1 | pandemic of "swine" flu [now designated A(H1N1)pdm09] |
Until 2009, these data suggest that flu pandemics occur when the virus acquires a new hemagglutinin and/or neuraminidase. For this reason, when an H1N1 virus appeared in a few recruits at Fort Dix in New Jersey in 1976, it triggered a massive immunization program (which turned out not to be needed). However, an H1N1 virus appeared the following year (perhaps escaped from a laboratory) causing the "Russian" flu. We now know that this virus was a direct descendant of the 1918 flu. While accumulating mutations that made it less dangerous, it had been infecting humans until it was replaced by the H2N2 "Asian" flu of 1957. Because most people born before the Asian flu pandemic of 1957 had been exposed to the H1N1 viruses circulating before, the Russian flu primarily affected children and young adults. For the same reason, this pattern was also seen in the 2009-10 pandemic of "swine" flu.
Where do the new H or N molecules come from?
Birds appear to be the source. Both the H2 that appeared in 1957 and the H3 that appeared in 1968 came from influenza viruses circulating in birds. The encoding of H and N by separate RNA molecules probably facilitates the reassortment of these genes in animals simultaneously infected by two different subtypes. For example, H3N1 virus has been recovered from pigs simultaneously infected with swine flu virus (H1N1) and the Hong King virus (H3N2). Probably reassortment can also occur in humans with dual infections.
Epidemics and Antigenic Drift
No antigenic shifts occurred between 1957 ("Asian") and 1968 ("Hong Kong"). So what accounts for the epidemics of 1962 and 1964? Missense mutations in the hemagglutinin (H) gene. Flu infections create a strong antibody response. After a pandemic or major epidemic, most people will be immune to the virus strain that caused it. The flu virus has two options:
- wait until a new crop of susceptible young people comes along
- change the epitopes on the hemagglutinin molecule (and, to a lesser degree, the neuraminidase) so that they are no longer recognized by the antibodies circulating in the bodies of previous victims.
- By 1972, the H3 molecules of the circulating strains differed in 18 amino acids from the original "Hong Kong" strain
- By 1975, the difference had increased to 29 amino acids.
The gradual accumulation of new epitopes on the H (and N) molecules of flu viruses is called antigenic drift. Spontaneous mutations in the H (or N) gene give their owners a selective advantage as the host population becomes increasingly immune to the earlier strains.
Flu Vaccines
Although a case of the flu elicits a strong immune response against the strain that caused it, the speed with which new strains arise by antigenic drift soon leaves one susceptible to a new infection. Immunization with flu vaccines has proved moderately helpful in reducing the size and severity of new epidemics.
Some vaccines incorporate inactivated virus particles; others use the purified hemagglutinin and neuraminidase. Both types incorporate antigens from the three major strains in circulation, currently:
- an A strain of the H1N1 subtype
- an A strain of the H3N2 subtype and
- a B strain.
Because of antigenic drift, the strains used must be changed periodically as new strains emerge that are no longer controlled by people's residual immunity.
The process:
- Chicken eggs are infected with the virus expressing the new H and/or N and simultaneously infected with a stock flu virus that grows very well in eggs.
- Genetic reassortment produces some viruses with both the new H and N genes along with the 8 other genes from the stock strain.
- This new virus is then grown in massive amounts and the H and N proteins purified for the new vaccine.
The whole process takes several weeks. A promising way to speed things up is to chemically synthesize the new H and N genes and substitute them for the H and N genes in the stock virus. The new virus can be ready for vaccine production in a few days.
| Season | H1N1 | H3N2 | Type B |
|---|---|---|---|
| 86–87 | A/Chile/83* | A/Mississippi/85 | B/Ann Arbor/86 |
| * As the 86–87 season got underway, it was found that A/Chile/83 no longer gave protection so A/Taiwan/86 was offered as a second shot late in that season. | |||
| 87–88 | A/Taiwan/86 | A/Leningrad/86 | B/Ann Arbor/86 |
| 88–89 | A/Taiwan/86 | A/Sichuan/87 | B/Victoria/87 |
| 89–90 | A/Taiwan/86 | A/Shanghai/87 | B/Yamagata/88 |
| 90–91 | A/Taiwan/86 | A/Shanghai/89 | B/Yamagata/88 |
| 91–92 | A/Taiwan/86 | A/Beijing/89 | B/Panama/90 |
| 92–93 | A/Texas/91 | A/Beijing/89 | B/Panama/90 |
| 93–94 | unchanged | unchanged | unchanged |
| 94–95 | A/Texas/91 | A/Shandong/93 | B/Panama/90 |
| 95–96 | A/Texas/91 | A/Johannesburg/94 | B/Harbin/94 |
| 96–97 | A/Texas/91 | A/Nanchang/95 | B/Harbin/94 |
| 97–98 | A/Johannesburg/96 | A/Nanchang/95 | B/Harbin/94 |
| 98–99 | A/Beijing/95 | A/Sydney/97 | B/Beijing/93 |
| 99–00 | A/Beijing/95 | A/Sydney/97 | B/Yamanashi/98 |
| 00–01 | A/New Caledonia/99 | A/Panama/99 | B/Yamanashi/98 |
| 01–02 | A/New Caledonia/99 | A/Panama/99 | B/Victoria/00 or similar |
| 02–03 | A/New Caledonia/99 | A/Moscow/99 | B/Hong Kong/2001 |
| 03–04 | A/New Caledonia/99 | A/Moscow/99 | B/Hong Kong/2001 |
| 04–05 | A/New Caledonia/99 | A/Fujian/2002 | B/Shanghai/2002 |
| 05–06 | A/New Caledonia/99 | A/California/2004 | B/Shanghai/2002 |
| 06–07 | A/New Caledonia/99 | A/Wisconsin/2005 | B/Malaysia/2004 |
| 07–08 | A/Solomon Islands/06 | A/Wisconsin/2005 | B/Malaysia/2004 |
| The B/Malasia component of the vaccine provided no protection at all. So all three components of the 08–09 vaccine were changed as shown on the next line. | |||
| 08–09 | A/Brisbane/2007 | A/Brisbane/2007 | B/Florida/2006 |
| 09–10 | A/Brisbane/2007 | A/Brisbane/2007 | B/Brisbane/2008 |
| Because the 2009–2010 pandemic of the newly-emerged "swine flu" virus drove the "seasonal" H1N1 viruses (e.g., A/Brisbane/2007) to near extinction, the "swine flu" H1N1 – now called A(H1N1)pdm09 – replaced the "seasonal" H1N1 in the 10–11 vaccine. |
|||
| 10–11 | A/California/2009 | A/Perth/2009 | B/Brisbane/2008 |
| 11–12 | All three components were unchanged from the previous year | ||
| 12–13 | A/California/2009 | A/Victoria/2011 | B/Wisconsin/2010 |
| 13–14 | A/California/2009 | A/Victoria/2011 | B/Massachusetts/2012 |
| 14–15 | A/California/2009 | A/Texas/2012 | B/Massachusetts/2012 |
| 15–16 | A/California/2009 | A/Switzerland/2013 | B/Phuket/2013 |
| 16–17 | A/California/2009 | A/HongKong/2014 | B/Brisbane/2008 |
| Several vaccines for the 2016-17 season will be quadrivalent; that is, contain a fourth component B/Phuket/2013. | |||
FluMist®
On 17 June 2003, the U. S. Food and Drug Administration (FDA) approved FluMist® – a live-virus vaccine that is given as a spray up the nose. The viruses have been weakened so that they do not cause illness, but are able to replicate in the relatively cool tissues of the nasopharynx where they can induce an immune response. Presumably this is tilted towards IgA production, a better defense against infection by inhaled viruses than blood-borne IgG antibodies. In any case, FluMist® induces a more rapid response than inactivated vaccine and there is some evidence that it provides better protection against antigenic drift as well.
All three currently-circulating strains of flu (H1N1, H3N2, and B) are included. As new strains appear, they can be substituted.
At present, this new vaccine (technically known as LAIV "Live Attenuated Influenza Vaccine") is only approved for children older than 24 months and adults younger than 50. People with immunodeficiency (e.g., AIDS) should also be cautious about taking it.
Update: For as yet unknown reasons, the nasal spray did not work during the 2015–2016 season, and it is not recommended for the upcoming season.
Flublok®
On 16 January 2013, the U. S. FDA approved an entirely new type of vaccine. Flublok® is made in cell cultures transformed with recombinant DNA encoding the hemagglutinins of the 3 currently circulating flu strains (H1N1, H3N2, and B). The final concentration of antigens is three times that in the current vaccine. Cultures of insect cells are used so there is no problem with possible egg allergies in those receiving the vaccine.
Other weapons against flu
It takes a while for the flu vaccine to build up a protective level of antibodies. What if you neglected to get your flu shot and now an epidemic has arrived?
Amantadine and Rimantadine
These drugs inhibit the M2 matrix protein needed to get viral RNA into the cytosol. They work against A strains only, and resistance to the drugs evolves quickly. By the 2009-2010 flu season, virtually all strains of both H3N2 and H1N1 had developed resistance.
Zanamivir (Relenza®) and Oseltamivir (Tamiflu®)
These drugs block the neuraminidase and thus inhibit the release and spread of fresh virions. Spraying zanamivir into the nose or inhaling it shortens the duration of disease symptoms by one to three days. Unfortunately, by the 2008-2009 flu season, all H1N1 strains circulating in the U.S. had become resistant to Tamiflu.
Antibiotics
Antibiotics are of absolutely no value against the flu virus. However, they are often given to patients to combat the secondary bacterial infections that occur and that are usually the main cause of serious illness and death.
Why so few drugs?
The mechanisms by which amantadine and zanamivir work provide a clue. There are far fewer anti-viral drugs than antibacterial drugs because so much of the virus life cycle is dependent on the machinery of its host. There are many agents that could kill off the virus, but they would kill off host cell as well. So the goal is to find drugs that target molecular machinery unique to the virus. The more we learn about these molecular details, the better the chance for developing a successful new drug.
The "Spanish" Flu
Jeffery Taubenberger and his colleagues have sequenced the genes of the influenza virus that had been recovered from
- preserved lung tissue of a U.S. soldier who died from influenza in 1918
- lung tissue from a flu victim whose body had remained frozen in the permafrost of Alaska since she died in 1918
But even with all of its genes now completely sequenced, why the 1918 strain was so deadly is not fully understood. But deadly it is. They have even been able to replace the 8 genes of a laboratory strain of flu virus with all 8 genes of the 1918 strain (using strict biosafety containment procedures!). The resulting virus kills mice faster than any other human flu virus tested. (Reported in the 7 October 2005 issue of Science.)
The Swine Flu of 2009
A new H1N1 flu began infecting humans in North America in April 2009 and has now spread throughout much of the world. Sequencing its genome revealed a novel virus - now called A(H1N1)pdm09 - that contained genes previously found in four different strains of swine flu:
- an HA gene (H1) derived from the swine flu of 1930 (and closely-related to the H1 of the great 1918 "Spanish" flu pandemic) along with an NP and NS gene from that virus;
- an NA gene (N1) from a virus that had been circulating in the pigs of Europe and Asia since 1979 along with the M gene from that virus;
- a PA and PB2 gene that entered pigs from birds around 1998;
- a PB1 gene that passed from birds to humans around 1968 and from us to pigs around 1998.
Why this remarkable assortment of genes has enabled he virus to jump so successfully from pigs to humans remains to be determined.
The amino acid sequence around the critical epitopes of its H1 molecules closely resemble those found in the resurrected 1918 flu virus. This would explain why
- Antibodies from elderly survivors of the 1918 pandemic neutralize the new swine flu virus.
- Antibodies (raised in mice) to the new swine flu virus neutralize the resurrected 1918 flu virus.
- The recent pandemic caused serious illness and death mostly in young adults and least in children and the elderly. As for the elderly, this contrast to the usual pattern arose because people over 65, even if not old enough to have been exposed to the 1918 virus, had been exposed to H1 viruses that until 1957 had only drifted from the original 1918 virus, and thus they had developed partial immunity. The antibodies in young adults were specific for seasonal flu strains circulating since 1957. These were unable to protect them against the 2009 virus but may have formed damaging immune complexes with them. Youngsters had no anti-flu antibodies and did not form such immune complexes.
"Bird Flu"
Many influenza A viruses are found in birds, both domestic and wild. Most of these cause little or no illness in these hosts. However, some of their genes can enter viruses able to infect domestic animals, as was the case for the PA and PB2 genes of the swine flu of 2009 (above).
On several occasions, bird flu viruses have also infected humans, often with alarmingly-high fatality rates. In 2003, human cases of an H7N7 bird flu virus infection occurred in the Netherlands, and in the same year an H5N1 bird virus caused human cases in large areas of Asia. Most of the human cases seemed to have been acquired from contact with infected birds rather than from human-to-human transmission.
And now in 2013, a new bird flu virus, H7N9, has appeared in humans in China. By the end of the summer of 2013, it had caused 135 observed cases (no one knows yet whether there may also be infected people who are not sick enough to show up at hospitals). 45 of the observed cases were fatal. The victims appear to have been infected through contact with infected poultry with little or no evidence of human-to-human transmission.
As a glance at the tables above will show, humans have had long experience with infections and vaccines by both H1 and H3 flu viruses. But the human population has absolutely no immunity against any H7 viruses. If this virus develops the capability to spread efficiently from human to human, it could lead to another worldwide pandemic.
Contributors and Attributions
John W. Kimball. This content is distributed under a Creative Commons Attribution 3.0 Unported (CC BY 3.0) license and made possible by funding from The Saylor Foundation.