
17.1: Parasitism
- Page ID
- 92879

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)
A fish parasite, the isopod Cymothoa exigua, replacing the tongue of a Lithognathus
Parasitism is a close relationship between species, where one organism, the parasite, lives on or inside another organism, the host, causing it some harm, and is adapted structurally to this way of life (Poulin 2007). Like predation, parasitism is a type of consumer-resource interaction (Getz 2011), but unlike predators, parasites, with the exception of parasitoids, are typically much smaller than their hosts, do not kill them, and often live in or on their hosts for an extended period. Parasites of animals are highly specialized, and reproduce at a faster rate than their hosts. Classic examples include interactions between vertebrate hosts and tapeworms, flukes, the malaria-causing Plasmodium species, and fleas.
Parasites reduce host fitness by general or specialized pathology, from parasitic castration to modification of host behavior. Parasites increase their own fitness by exploiting hosts for resources necessary for their survival, in particular by feeding on them and by using intermediate (secondary) hosts to assist in their transmission from one definitive (primary) host to another. Although parasitism is often unambiguous, it is part of a spectrum of interactions between species, grading via parasitoidism into predation, through evolution into mutualism, and in some fungi, shading into being saprophytic.
Within that scope are many possible strategies. Taxonomists classify parasites in a variety of overlapping schemes, based on their interactions with their hosts and on their life-cycles, which are sometimes very complex. An obligate parasite depends completely on the host to complete its life cycle, while a facultative parasite does no. Parasite life-cycles involving only one host are called "direct"; those with a definitive host (where the parasite reproduces sexually) and at least one intermediate host are called "indirect" (Garcia 1999). An endoparasite lives inside the host's body; an ectoparasite lives outside, on the host's surface (Australian Society of Parasitology 2010). Mesoparasites—like some copepods, for example—enter an opening in the host's body and remain partly embedded there (Vecchione and Aznar 2008). Some parasites can be generalists, feeding on a wide range of hosts, but many parasites, and the majority of protozoans and helminths that parasitise animals, are specialists and extremely host-specific (Australian Society of Parasitology 2010). The microorganisms and viruses that can reproduce and complete their life cycle within the host are known as microparasites. Macroparasites are the multicellular organisms that reproduce and complete their life cycle outside of the host or on the host's body (Poulin 2011; Blackwell Science 2018).
Strategies
There are six major parasitic strategies, namely parasitic castration; directly transmitted parasitism; trophically-transmitted parasitism; vector-transmitted parasitism; parasitoidism; and micropredation. These apply to parasites whose hosts are plants as well as animals (Poulin 2011; Poulin 2015). These strategies represent adaptive peaks; intermediate strategies are possible, but organisms in many different groups have consistently converged on these six, which are evolutionarily stable (Poulin 2011).
A perspective on the evolutionary options can be gained by considering four key questions: the effect on the fitness of a parasite's hosts; the number of hosts they have per life stage; whether the host is prevented from reproducing; and whether the effect depends on intensity (number of parasites per host). From this analysis, the major evolutionary strategies of parasitism emerge, alongside predation (Lafferty and Kuris 2002).
|
Host fitness |
Single host, stays alive |
Single host, dies |
Multiple hosts |
|
Able to reproduce (fitness > 0) |
Conventional parasite Pathogen |
Trophically-transmitted parasite Trophically-transmitted pathogen |
Micropredator Micropredator |
|
Unable to reproduce (fitness = 0) |
----- Parasitic castrator |
Trophically-transmitted parasitic castrator Parasitoid |
Social predator Solitary predator |
Parasitic castrators

The parasitic castrator Sacculina carcini (highlighted) attached to its crab host
Parasitic castrators partly or completely destroy their host's ability to reproduce, diverting the energy that would have gone into reproduction into host and parasite growth, sometimes causing gigantism in the host. The host's other systems remain intact, allowing it to survive and to sustain the parasite (Poulin 2015; Poulin 2007). Parasitic crustaceans such as those in the specialized barnacle genus Sacculina specifically cause damage to the gonads of their many species (Elumalai et al. 2013) of host crabs. In the case of Sacculina, the testes of over two-thirds of their crab hosts degenerate sufficiently for these male crabs to develop female secondary sex characteristics such as broader abdomens, smaller claws and egg-grasping appendages. Various species of helminth castrate their hosts (such as insects and snails). This may happen directly, whether mechanically by feeding on their gonads, or by secreting a chemical that destroys reproductive cells; or indirectly, whether by secreting a hormone or by diverting nutrients. For example, the trematode Zoogonus lasius, whose sporocysts lack mouths, castrates the intertidal marine snail Tritia obsoleta chemically, developing in its gonad and killing its reproductive cells (Poulin 2007; Cheng 2012).
Directly-Transmitted

Human head-lice exemplify directly transmitted obligate ectoparasites
Directly-transmitted parasites, not requiring a vector to reach their hosts, include such parasites of terrestrial vertebrates as lice and mites; marine parasites such as copepods and cyamid amphipods; monogeneans; and many species of nematodes, fungi, protozoans, bacteria, and viruses. Whether endoparasites or ectoparasites, each has a single host-species. Within that species, most individuals are free or almost free of parasites, while a minority carry a large number of parasites; this is known as an aggregated distribution (Poulin 2015).
Trophically-transmitted
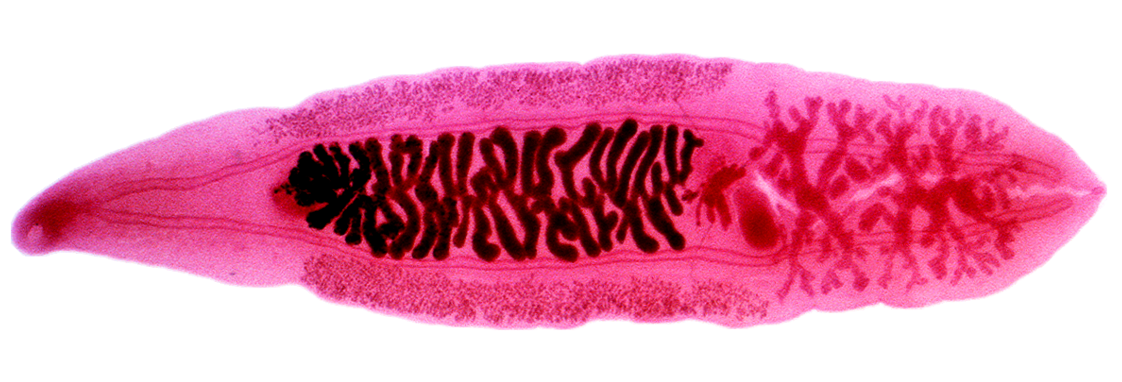
Clonorchis sinensis, the Chinese liver fluke, is trophically transmitted
Trophically-transmitted parasites are transmitted by being eaten by a host. They include trematodes (all except schistosomes), cestodes, acanthocephalans, pentastomids, many round worms, and many protozoa such as Toxoplasma (Poulin 2015). They have complex life-cycles involving hosts of two or more species. In their juvenile stages they infect and often encyst in the intermediate host. When the intermediate-host animal is eaten by a predator, the definitive host, the parasite survives the digestion process and matures into an adult; some live as intestinal parasites. Many trophically-transmitted parasites modify the behavior of their intermediate hosts, increasing their chances of being eaten by a predator. As with directly transmitted parasites, the distribution of trophically transmitted parasites among host individuals is aggregated (Poulin 2015). Coinfection by multiple parasites is common (Cox 2001). Autoinfection, where (by exception) the whole of the parasite's life-cycle takes place in a single primary host, can sometimes occur in helminths such as Strongyloides stercoralis (Australian Society of Parasitology 2017).
Vector-transmitted
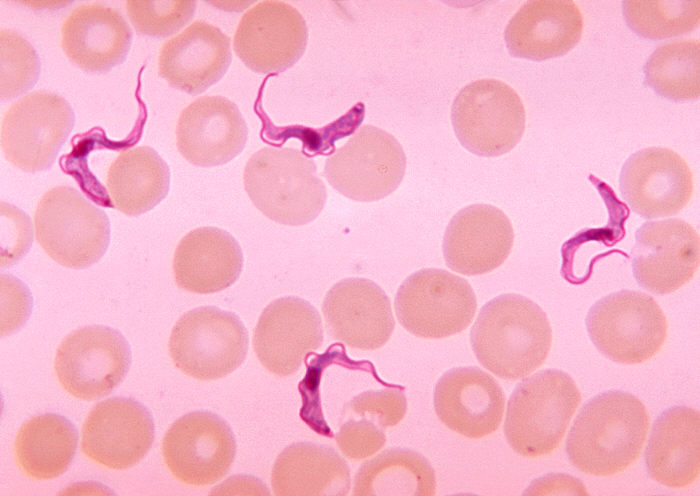
The vector-transmitted protozoan endoparasite Trypanosoma among human red blood cells
Vector-transmitted parasites rely on a third party, an intermediate host, where the parasite does not reproduce sexually (Australian Society of Parasitology 2010) to carry them from one definitive host to another (Poulin and Randhawa 2015). These parasites are microorganisms, namely protozoa, bacteria, or viruses, often intracellular pathogens (disease-causers) (Poulin and Randhawa 2015). Their vectors are mostly hematophagic arthropods such as fleas, lice, ticks, and mosquitoes (Poulin and Randhawa 2015, PEOI 2013). For example, the deer tick Ixodes scapularis acts as a vector for diseases including Lyme disease, babesiosis, and anaplasmosis (Steere 2001). Protozoan endoparasites, such as the malarial parasites in the genus Plasmodium and sleeping-sickness parasites in the genus Trypanosoma, have infective stages in the host's blood which are transported to new hosts by biting insects (Pollitt et al. 2011).
Parasitoids
Parasitoids are insects which sooner or later kill their hosts, placing their relationship close to predation (Stevens 2010). Most parasitoids are parasitoid wasps or other hymenopterans; others include dipterans such as phorid flies. They can be divided into two groups, idiobionts and koinobionts, differing in their treatment of their hosts (Gullan and Cranston 2010).
Idiobiont parasitoids sting their often large prey on capture, either killing them outright or paralyzing them immediately. The immobilized prey is then carried to a nest, sometimes alongside other prey if it is not large enough to support a parasitoid throughout its development. An egg is laid on top of the prey and the nest is then sealed. The parasitoid develops rapidly through its larval and pupal stages, feeding on the provisions left for it (Gullan and Cranston 2010).
Idiobiont parasitoid wasps immediately paralyze their hosts for their larvae (Pimplinae, pictured) to eat (Poulin and Randhawa 2015).

Koinobiont parasitoids, which include flies as well as wasps, lay their eggs inside young hosts, usually larvae. These are allowed to go on growing, so the host and parasitoid develop together for an extended period, ending when the parasitoids emerge as adults, leaving the prey dead, eaten from inside. Some koinobionts regulate their host's development, for example preventing it from pupating or making it molt whenever the parasitoid is ready to molt. They may do this by producing hormones that mimic the host's molting hormones (ecdysteroids), or by regulating the host's endocrine system (Gullan and Cranston 2010).
Koinobiont parasitoid wasps like this braconid lay their eggs inside their hosts, which continue to grow and moult.

Phorid fly (center left) is laying eggs in the abdomen of a worker honey-bee, altering its behavior.

Micropredators
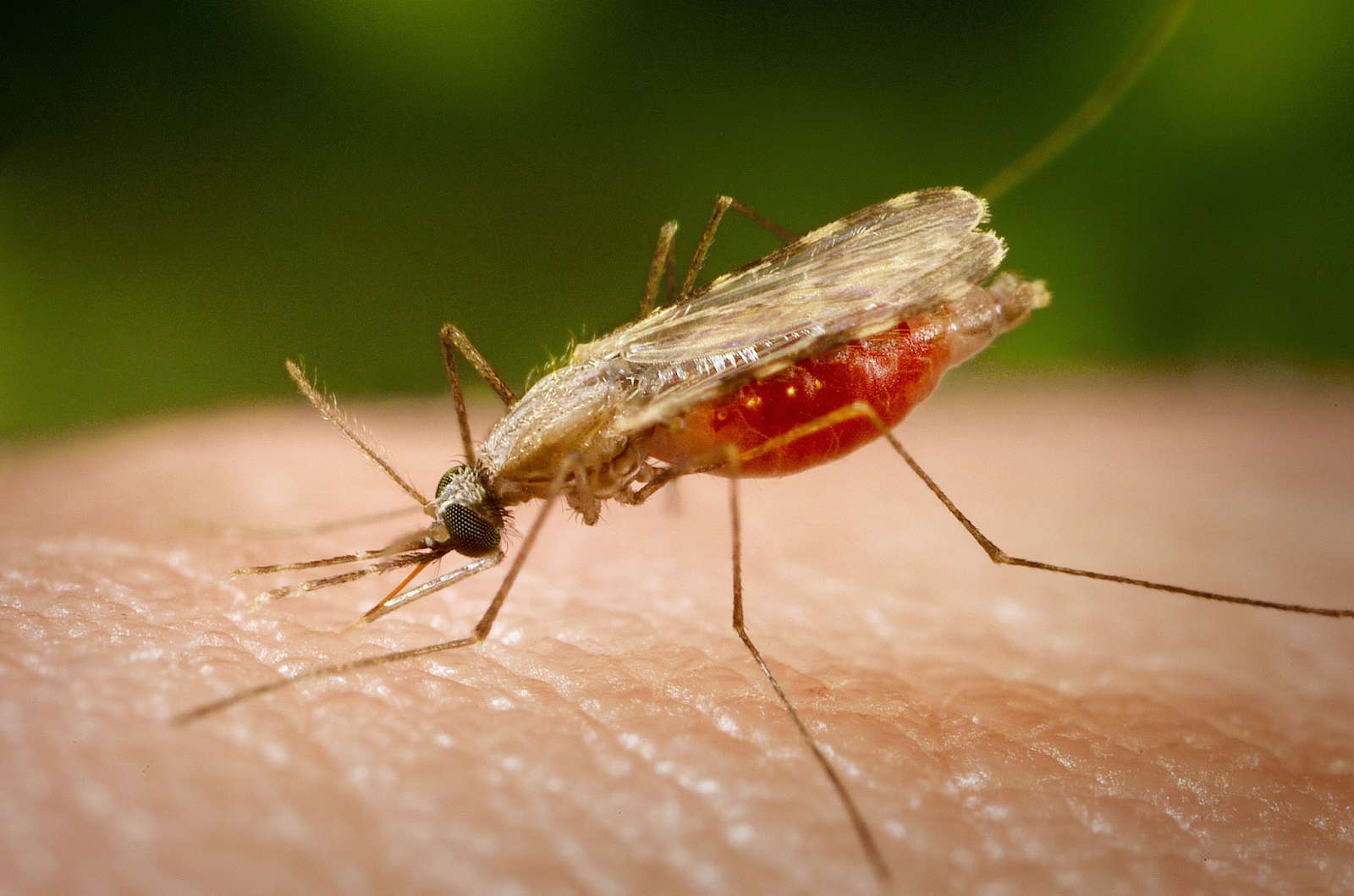
Mosquitoes are micropredators, and important vectors of disease
A micropredator attacks more than one host, reducing each host's fitness by at least a small amount, and is only in contact with any one host intermittently. This behavior makes micropredators suitable as vectors, as they can pass smaller parasites from one host to another (Poulin and Randhawa 2015; Lafferty and Kuris 2002; Wilson et al. 2017). Most micropredators are hematophagic, feeding on blood. They include annelids such as leeches, crustaceans such as branchiurans and gnathiid isopods, various dipterans such as mosquitoes and tsetse flies, other arthropods such as fleas and ticks, vertebrates such as lampreys, and mammals such as vampire bats (Poulin and Randhawa 2015).
Variations on Parasitism
Hyperparasitism
Hyperparasites feed on another parasite, as exemplified by protozoa living in helminth parasites (Dissanaike 1957), or facultative or obligate parasitoids whose hosts are either conventional parasites or parasitoids (Poulin and Randhawa 2015; Gullan and Cranston 2010). Levels of parasitism beyond secondary also occur, especially among facultative parasitoids. In oak gall systems, there can be up to five levels of parasitism (Askew 1951).
Hyperparasites can control their hosts' populations, and are used for this purpose in agriculture and to some extent in medicine. The controlling effects can be seen in the way that the CHV1 virus helps to control the damage that chestnut blight, Cryphonectria parasitica, does to American chestnut trees, and in the way that bacteriophages can limit bacterial infections. It is likely, though little researched, that most pathogenic microparasites have hyperparasites which may prove widely useful in both agriculture and medicine (Parratt and Laine 2016).
A hyperparasitoid pteromalid wasp on the cocoons of its host, itself a parasitoid braconid wasp

Social parasitism
Social parasites take advantage of interspecific interactions between members of eusocial animals such as ants, termites, and bumblebees. Examples include the large blue butterfly, Phengaris arion, its larvae employing ant mimicry to parasitise certain ants (Thomas et al. 2010), Bombus bohemicus, a bumblebee which invades the hives of other bees and takes over reproduction while their young are raised by host workers, and Melipona scutellaris, a eusocial bee whose virgin queens escape killer workers and invade another colony without a queen (Van Oystaeyen et al. 2013) An extreme example of interspecific social parasitism is found in the ant Tetramorium inquilinum, an obligate parasite which lives exclusively on the backs of other Tetramorium ants (Antkeepers 2016).
The large blue butterfly is an ant mimic and social parasite.

Brood parasitism
In brood parasitism, the hosts act as parents as they raise the young as their own. Brood parasites include birds in different families such as cowbirds, whydahs, cuckoos, and black-headed ducks. These do not build nests of their own, but leave their eggs in nests of other species. The eggs of some brood parasites mimic those of their hosts, while some cowbird eggs have tough shells, making them hard for the hosts to kill by piercing, both mechanisms implying selection by the hosts against parasitic eggs (Payne 1997; Rothstein 1990; De Marsico et al. 2013). The adult female European cuckoo further mimics a predator, the European sparrowhawk, giving her time to lay her eggs in the host's nest unobserved (Welbergen and Davies 2011).
In brood parasitism, the host raises the young of another species, here a cowbird's egg, that has been laid in its nest.

Kleptoparasitism
In kleptoparasitism (from Greek κλέπτης (kleptēs), "thief"), parasites steal food gathered by the host. The parasitism is often on close relatives, whether within the same species or between species in the same genus or family. For instance, the many lineages of cuckoo bees lay their eggs in the nest cells of other bees in the same family (Slater et al. 2005). Kleptoparasitism is uncommon generally but conspicuous in birds; some such as skuas are specialized in pirating food from other seabirds, relentlessly chasing them down until they disgorge their catch (Furness 1978).
The great skua is a powerful kleptoparasite, relentlessly pursuing other seabirds until they disgorge their catches of food.

Sexual parasitism
A unique approach is seen in some species of anglerfish, such as Ceratias holboelli, where the males are reduced to tiny sexual parasites, wholly dependent on females of their own species for survival, permanently attached below the female's body, and unable to fend for themselves. The female nourishes the male and protects him from predators, while the male gives nothing back except the sperm that the female needs to produce the next generation (Pietsch 2005).
The male anglerfish Ceratias holboelli lives as a tiny sexual parasite permanently attached below the female's body.

Adelphoparasitism
Adelphoparasitism, (from Greek ἀδελφός (adelphós), brother – Maggenti et al. 2005), also known as sibling-parasitism, occurs where the host species is closely related to the parasite, often in the same family or genus (Rochat and Gutierrez 2001). In the citrus blackfly parasitoid, Encarsia perplexa, unmated females of which may lay haploid eggs in the fully developed larvae of their own species, producing male offspring (University of Florida 2018), while the marine worm Bonellia viridis has a similar reproductive strategy, although the larvae are planktonic (Berec et al. 2005).
Encarsia perplexa (center), a parasitoid of citrus blackfly (lower left), is also an adelphoparasite, laying eggs in larvae of its own species
