3.2: Enzymes
- Page ID
- 64247

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Biological catalysts are called enzymes, and the overwhelming majority of enzymes are proteins. The exceptions are a class of RNA molecules known as ribozymes, of which most act upon themselves (i.e. part of the RNA strand is a substrate for the ribozyme part of the strand). In this book (and most textbooks in this field), unless otherwise specified, the term enzyme refers to one made of protein. Enzymes confer extraordinary specificity to a chemical reaction: a reaction that might occur between a variety of potential substrates in an uncatalyzed situation may only be allowed between two specific substrates when catalyzed by an enzyme. Enzymes allow cells to run chemical reactions at rates from a million to even a trillion times faster than the same reactions would run under similar conditions without enzymes. In some cases, the enzymes al - low reactions to proceed that would normally (i.e. sans enzyme) require more extreme temperature, pressure, or acidity/alkalinity. Finally, and perhaps most importantly for life, enzymes can be regulated. This is crucial for the cell, since it must be able to react to different situations, such as availability of energy, accumulation of toxic byproducts, the need to reproduce, etc. Not only can enzymes be modified either covalently or noncovalently to increase or decrease their activity, the cell can also regulate production of the enzymes, providing another level of control over particular cellular biochemical reactions.
Enzymes have been cataloged and classified since the 1950’s, during which time there was an explosion of enzyme discoveries and a need for a unified nomenclature and catalog. An International Commission on Enzymes was established and thus started the Enzyme List. All enzymes now have both recommended names for common usage, often reflecting historical naming, and a systematic name, which is highly specific. They also have a classification number based on their activity. The major classes of enzymes are
- Oxidoreductases that carry out oxidation-reduction reactions,
- Transferases that transfer functional groups,
- Hydrolases that carry out hydrolysis reactions,
- Lyases that eliminate groups to form double bonds,
- Isomerases that rearrange the bonds in a molecule but do not add or remove atoms, and
- Ligases that form bonds in reactions coupled to ATP hydrolysis.
As an example, DNA ligase (recommended name) catalyzes the formation of a phosphodiester bond between the 3’ end of one DNA fragment and the 5’ end of another. Its rather long and tedious systematic name is “poly(deoxyribonucleotide):poly(deoxyr ibonucleotide) ligase (AMP-forming)” and its classification number is 6.5.1.1. As a ligase, it is class 6; because it forms phosphoric ester bonds, it is subclass 5; the sub-subclass of 1 in this case is meaningless because it is the only sub-subclass of phosphoricester bond-forming ligases, but the final number designates the DNA ligase separately from other 6.5.1 enzymes such as RNA ligase, which is 6.5.1.3.
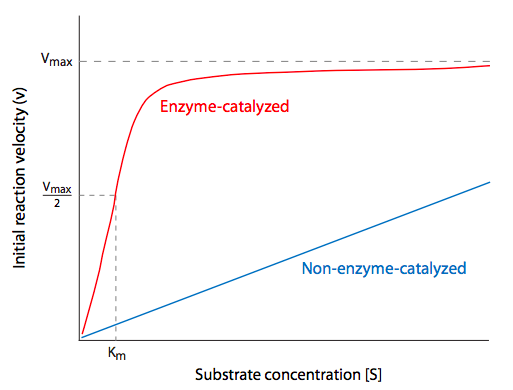
Enzymes are the most diverse type of protein in a cell. They vary not only in size, but also in the number of independently manufactured subunits that must come together to form an active enzyme, or holoenzyme. Part of the reason for requiring so many different enzymes is that they are usually very specific for their substrate molecules, and that specificity is based upon a combination of shape and charge. The interactions between substrate and enzyme are often likened to a lock and key or pieces of a jigsaw puzzle. If the substrate fits the shape of the enzyme’s active site (the part of the enzyme that carries out the actual catalytic reaction), and the charges interact (e.g. positively charged amino acids on the enzyme lining up with negative charges on the substrate), then there may be further stabilization of the interaction by Van der Waals and hydrogen bond interactions. In fact, formation of a stable Enzyme-Substrate (ES) intermediate is energetically analogous to the transition state (Figure \(\PageIndex{2}\)) of reactions.
The specificity of enzymes is such that stereoisomers may not be recognized by some enzymes: for example, a protease (enzymes that chop up proteins into smaller pieces by hydrolyzing the peptide bonds between specific amino acids) such as trypsin can be stymied by the presence of a D-amino acid in place of the usual L-amino acid in a protein, even though it is a mirror image of the very same amino acid. This specificity means that enzymes are highly selective with respect to the reactions they catalyze, which means that specific reactions can be greatly enhanced without causing a general increase in many related chemical reactions. Another implication of the high specificity is that enzymes can (and often do) have high affinity for their substrates without the problem of binding non-substrate molecules (other than specific inhibitors - see below).
If most biochemical reactions would proceed extremely slowly, if at all, without catalysis, enzymes are needed to lower the activation energy needed for chemical reactions to support life. Exactly how does an enzyme lower the activation energy of a reaction? What exactly does “activation energy” mean in the context of a cell? To understand this, there are two principles to keep in mind: first, when we talk about chemical reactions, generally, we are concerned with populations of substrate, product, and enzyme molecules, not individuals; and second, the reactions are generally taking place between molecules dissolved in the aqueous cytoplasm of the cell.
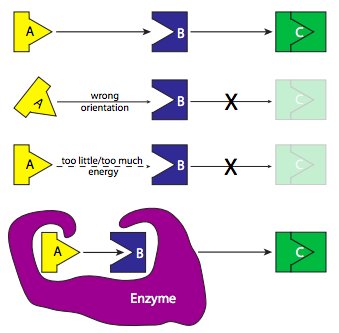
Consider a reaction in which substrates A and B interact to form product C (Figure \(\PageIndex{3}\)). If this reaction is not catalyzed, it depends on the happenstance that a molecule of A runs into a molecule of B in just the right orientation, and with the right amount of energy, to react and form the new molecule. We can conceptualize “activation energy” as the difficulty in getting A and B together perfectly so the reaction can proceed. How might an enzyme lower this activation energy? By making it easier for A and B to find each other with the right orientation and energy. So it could have binding sites for molecule A and molecule B, and once it has bound these two molecules, it changes its conformation, bringing A and B together under exactly the right conditions to react and form C. Once the reaction is complete, the product floats off because the enzyme has no affinity for it, and the enzyme returns to its initial shape, ready to bind more substrates.

Enzymes may also facilitate a chemical reaction by acting as a temporary holding site for an active group being transferred from one substrate to another. Alternatively, temporary formation of hydrogen bonds or even covalent bonds between the enzyme and substrate can alter chemical characteristics of the substrate to make it react more easily. An example of enzyme mechanisms on the molecular level is shown in Chapter 5: Figure \(\PageIndex{1}\).
Another example may be found with enzymes that break apart a molecule (Figure \(\PageIndex{4}\)). In order for a molecule to break apart, it may need to collide with another molecule with sufficient energy to break one or more of its covalent bonds. An enzyme that catalyzes the breakdown reaction might bind to the molecule, and in binding it, undergoes a conformational shift that bends or twists the molecule in such a way that the bonds in the substrate molecule are weakened or broken. These two examples oversimplify the chemistry of enzyme activity into a mechanical idea, but the general relationship in how an enzyme lowers activation energy for a reaction is accurate.