
3.8: Basic Nomenclature
- Page ID
- 42793

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Single Letter System
Sometimes what you want to do is a little rough work for investigating your genetic model. A genetic model is a diagram of the logic that you propose for a particular type of inheritance. For instance, if you cross a true‐breeding purple plant with a true‐breeding white plant (Figure A1.2), you will get a heterozygote (the middle plant). If we name the gene after the mutation (a is the first letter in “albino”), we know that the heterozygote will have one capital letter “A” and a lower case “a”. The heterozygote is the F1 generation (“first filial”, which means it’s the first child from parents that are crossed). The F1 is purple, which means the “a” allele is recessive; only one copy of the “A” allele is needed for enough purple pigment to make it identical to one true-breeding parent. This is complete dominance.
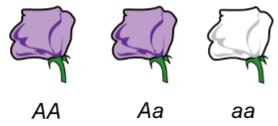
We cannot know from the information given which allele is wild type or mutant. One hypothesis, though, is that purple pigments are required to attract pollinators and therefore would help the plant in the wild. Albino plants could be a mutant and might not generate as many seeds for lack of pollinators. If the context of your genetics problem doesn’t indicate which allele is wild‐type, it’s good practice to name your allele based on the recessive trait. Often the recessive allele is the mutation. We might consider that the “a” allele is null and makes no pigment at all. One or two “A” alleles make enough protein to cause the plant to be purple.
We can’t know from the information given which allele is wild type or mutant. One hypothesis, though, is that purple pigments are required to attract pollinators and therefore would help the plant in the wild. Albino plants could be a mutant and might not generate as many seeds for lack of pollinators. If the context of your genetics problem doesn’t indicate which allele is wild‐type, it’s good practice to name your allele based on the recessive trait. Often the recessive allele is the mutation. We might consider that the “a” allele is null and makes no pigment at all. One or two “A” alleles make enough protein to cause the plant to be purple.
Figure A1.2 already assumes that the capital letter (A) stands for an allele that encodes a protein for purple pigment and the recessive allele (a) doesn’t make pigment. Thus the Aa heterozygote is sufficient evidence to adopt upper- and lower-case letter “A”s to communicate the characteristics of purple and white alleles.
A note of caution. When you’re writing down gene symbols for homework or on an exam, be sure to make the characters distinct. A typewritten “y” is easy to distinguish from the upper case “Y” but not as easy when writing it down. Instructors who ask you to show your work need to be able to follow your logic train. More important than that is YOU have to be able to follow your own reasoning. Students often switch symbols and come up with an answer that is inconsistent with the data given because of this. Consider underlining your capitals or putting a line through one of them to make it distinct (e.g. Y for the dominant allele; y for the recessive).
Name the gene after the mutant phenotype
Some instructors would accept “P” for “purple” for the previous cross. However, the better answer is to follow an established system. During “exam fog”, it’s easy to get lost if you are inconsistent with how you develop your symbols. During your study period and when you’re practicing genetics problems, be thoughtful about the gene names you choose.
Let’s always choose a letter based on the mutant phenotype for our gene symbol. If we are presented with a ladybug mutant that is small, we might choose “d” for “dwarf”. Geneticists sometimes set up a research program based on unusual phenotypes of the organism they are studying. The fact that a mutant phenotype that is heritable exists tells us that there is a genetic control for the trait and that it might be isolated in the lab. When you look at your classmates, you don’t necessarily note that none of them has an arm growing from the tops of their heads. If one student had this trait, however, you couldn’t help but notice it. If you found out it showed up in that student’s ancestry in a predictable fashion, you might reasonably suggest that there is a genetic basis for that. If it happened to be controlled by a single gene, you might call the gene “extra arm” or “arm head”. If it happened to be a dominant trait, you might use the letter “X” or “A” for the mutant allele. The wild type allele would be “x” or “a”, respectively.
you don’t know which is mutant, use the recessive trait for the gene name
What if you don’t know which allele is mutant? What if you’re presented with two true-‐ breeding frogs: one that is gold and one that is yellow. If you don’t know what the predominant colour in nature is you can’t know which one is mutant. If you crossed them and all the progeny are gold, then you know the dominant allele encodes a protein to make it gold. The recessive, therefore, is “yellow” and you should name the gene “y” after the recessive phenotype. This means the dominant allele would be “Y”. Your offspring would therefore be Yy and the gold parent would be YY. The yellow parent would be yy.
Apply these ideas at the Online Open Genetics exercises.
1.3 One Letter System
Superscripts
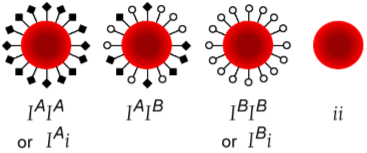
Sometimes a letter is used as the name of a gene, and superscripts can modify it to indicate the different alleles. One common single letter code for an allelic series is “I". Red blood cells can have their cell membranes modified by sugar tags that give rise to our blood type. One allele of I gives rise to blood type A and is therefore called IA. An enzyme encoded by IB modifies sugars to create blood type B. A heterozygote IAIB demonstrates both sugar tags because those alleles are expressed – they are codominant. People with blood type O only possess alleles for the I gene that don’t work and are therefore recessive – they don’t modify the extracellular sugar tags. Because it is recessive, individuals are homozygous for i: they are ii. Chapter 13, Section 7 has more detail on this allelic series.
Sometimes a superscript “plus sign” is used to denote the wild type allele. One might use the symbol W+ to indicate a wild-‐type allele that promotes wing growth. Note that the generic “wing” gene name isn’t a best practice – name the gene after the mutant phenotype! A wingless mutant would be W ‐. You should never use a “+” and shift the case of the letter unless you are dealing with a special case such as the codominance in the blood type example above. The capital “I” letter indicates it is dominant to “i". The superscript A and B for the codominant alleles indicate the dominant alleles are different from each other.
Superscripts can be symbols, a single letter, or many letters. They modify the gene name only in the superscripted symbols: the regular-‐sized letters are identical between them (see Table 1). This means that Abc+ and abc would be different genes (i.e. not allelic); Abc+ and Abc are alleles, as are abc+ and abc. Note that a superscript is not mandatory for all alleles of that gene, depending on the convention.
Alleles of bacterial genes are typically indicated with a superscript + or -. For example, a bacterial allele that creates an enzyme that makes methionine would be met+, and a defective allele of that gene is met -.
| whiteapr or whiteapricot | An allele of the white gene which has an “apricot” phenotype |
| Abc+ Abc | Two alleles for the Abc gene (wild-type and mutant, respectively). Note the mutant allele is dominant. |
| w+; wa; w | Three alleles in a series for the w gene. The first is wild type; the second two are different mutant alleles. |
| bio+; bio- | A wild-type allele of a biotin gene and its recessive counterpart. This is likely a bacterial gene due to the convention. |