
18.5: Macrofungi
- Page ID
- 123999

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Ascomycota
- Use life history traits and morphological features to distinguish between Ascomycota and Basidiomycota.
- Identify structures in the Ascomycota life cycle and know their ploidy.
- Differentiate between different types of ascocarps; locate fertile surfaces within those structures.
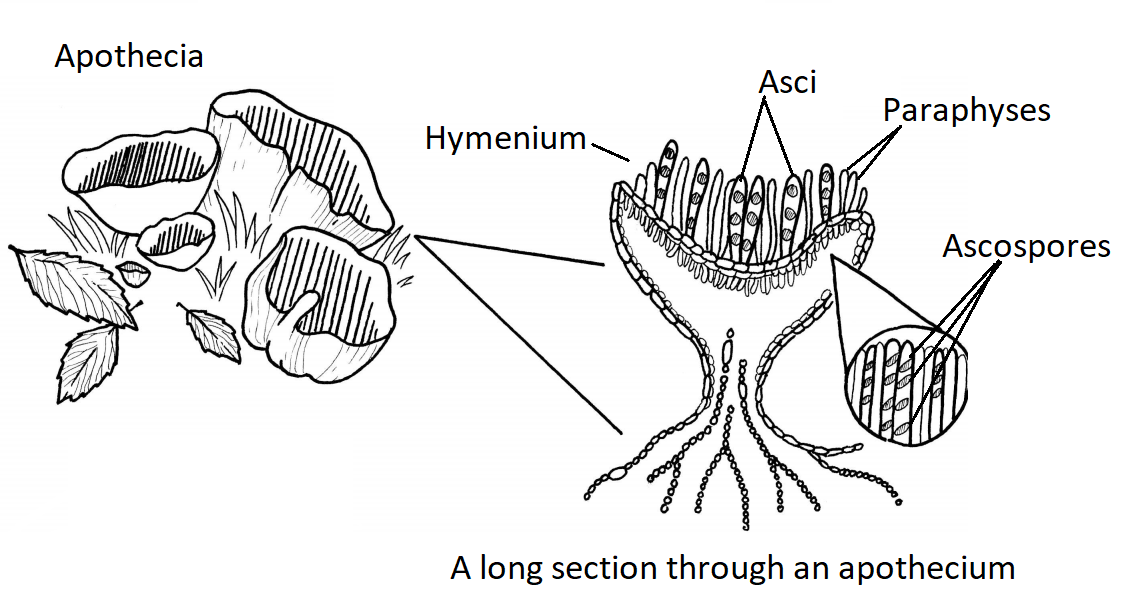
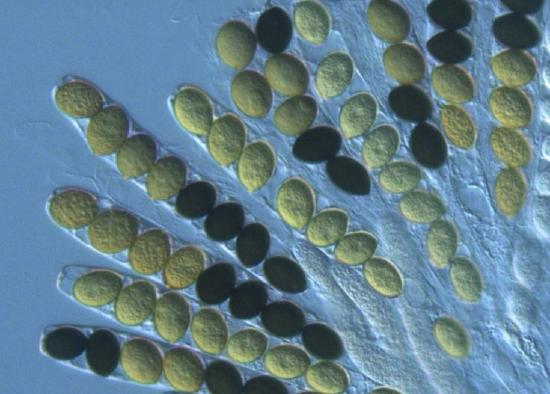
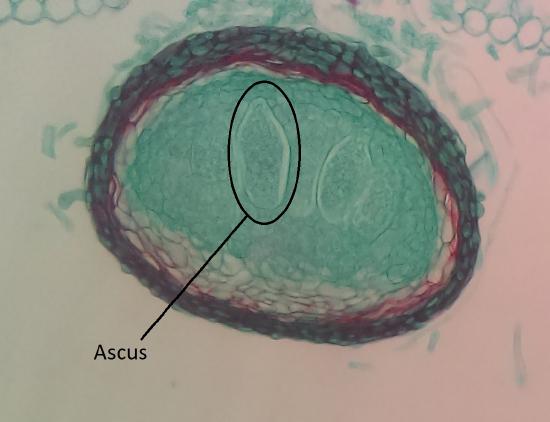
The majority of described fungal species belong to the Phylum Ascomycota. Fungi in this group have simple septate hyphae and most produce fruiting bodies called ascocarps (Figure \(\PageIndex{1}\)) for sexual reproduction. Some genera of Ascomycota either only reproduce asexually via conidia or the sexual phases have not been discovered or described. These were referred to as the Fungi Imperfecti or Deuteromycota. When ascomycetes reproduce sexually, they produce haploid ascospores (usually 8) within a sac-like structure called an ascus (Figure \(\PageIndex{2}\)).
The Ascomycota include some ectomycorrhizal fungi (e.g. truffles and morels), fungi that are used as food, others that are common causes of food spoilage (bread molds and plant pathogens), and still others that are human pathogens. Some notable examples of ascomycetes include:
- Saccharomyces cerevisiae one of the budding yeasts. It ferments sugar to ethanol and carbon dioxide and thus is used to make alcoholic beverages like beer and wine, to make ethanol for industrial use and in baking (it is often called baker's yeast). Here, it is the carbon dioxide that is wanted (to make bread and cakes "rise" and have a spongy texture). Yeast is also used in the commercial production of some vitamins and in the production - using recombinant DNA technology - of some human therapeutic proteins.
- Neurospora crassa, another favorite "model" organism in the laboratory.
- The fungal partner in most lichens is an ascomycete.
- Powdery mildews that attack ornamental plants.
- The chestnut blight, which in a few decades killed almost all of the mature American chestnut trees in the Appalachians of North America.
- The Dutch elm disease, which has killed many of the American elms in the United States.
- Pneumocystis jirovecii, which is a major cause of illness in immunosuppressed people, e.g., patients with AIDS.
- The truffle and the morel, both highly-prized food delicacies. Truffles establish a symbiotic relationship with the roots of such trees as oaks.

Ascomycete Life Cycle
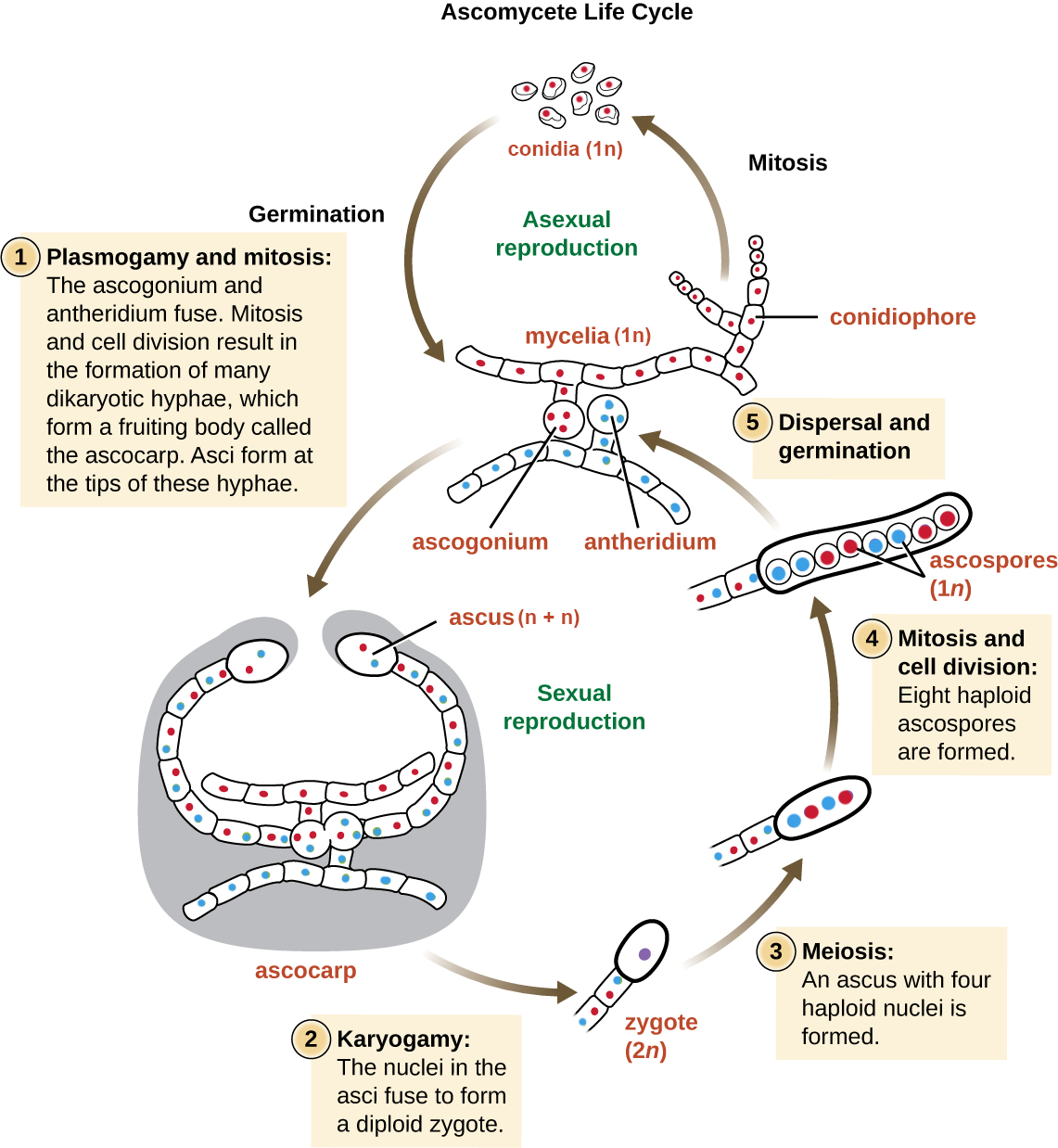
Asexual reproduction is frequent and involves the production of conidiophores that release haploid conidia. Sexual reproduction starts with the development of special hyphae from either one of two types of mating strains. One strain produces an antheridium and a strain of a complementary mating type develops an ascogonium. At fertilization, the antheridium and the ascogonium combine in plasmogamy without nuclear fusion. Dikaryotic ascogenous hyphae arise, in which pairs of nuclei migrate: one from each parent strain. In each ascus mother cell, two nuclei fuse (karyogamy). During sexual reproduction, thousands of asci may fill a fruiting body called the ascocarp. The diploid nucleus gives rise to four haploid nuclei by meiosis. In most ascomycetes, this is followed by a round of mitosis, producing 8 ascospores. The ascospores are then released, germinate, and form haploid hyphae that are disseminated in the environment and start new mycelia (Figure \(\PageIndex{4}\)).
Types of Ascocarps
Apothecium
Apothecia are cup-shaped with the asci fully exposed, lining the interior of the cup. Normally, these asci are microscopic. However, in the fungus Ascobolus, the large asci with dark ascospores can be seen with the naked eye or a handlens (Figure \(\PageIndex{5}\)). The typical cup shape can be inverted and take on strange morphologies (see Figure \(\PageIndex{6}\)).
.jpg?revision=1&size=bestfit&width=525&height=423)
.jpg?revision=1&size=bestfit&height=435) Figure \(\PageIndex{5}\): In the first photo, apothecia of Ascobolus furfuraceus have asci appear black due to their dark spores. In the second photo, a magnified view of Ascobolus immersus shows the dark ascospores. First photo by Salvatore Bacciu and Paola Mereu, (CC-BY-NC). Second photo by Jerry Cooper, some rights reserved (CC-BY).
Figure \(\PageIndex{5}\): In the first photo, apothecia of Ascobolus furfuraceus have asci appear black due to their dark spores. In the second photo, a magnified view of Ascobolus immersus shows the dark ascospores. First photo by Salvatore Bacciu and Paola Mereu, (CC-BY-NC). Second photo by Jerry Cooper, some rights reserved (CC-BY).
.jpeg?revision=1&size=bestfit&width=698&height=392)
Perithecium
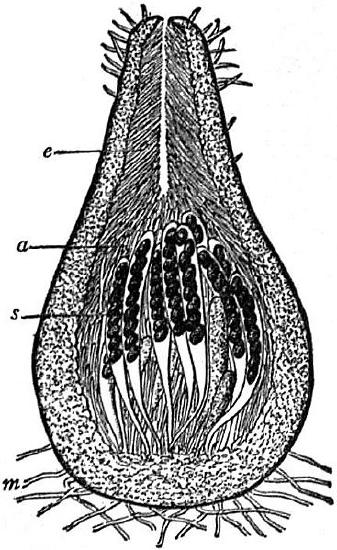
A perithecium is a flask-shaped fruiting structure (Figure \(\PageIndex{7}\)), often microscopic and embedded within either the substrate it is fruiting in or a fungal structure called a stroma (Figure \(\PageIndex{8}\)). The asci are almost entirely closed off from the external environment, excepting a small hole in the top of the perithecium called an ostiole. Many plant parasites, such as the causal agents of Dutch elm disease, American chestnut blight, and ergot, are perithecial ascomycetes.

.jpeg?revision=1&size=bestfit&width=317&height=476)
Cleistothecium
A cleistothecium is a fully-enclosed fruiting structure. These typically have bag-like asci (Figure \(\PageIndex{9}\)). Some (chasmothecia) split open to release their spores. This type of fruiting body can be found in the powdery mildews, a group of plant pathogenic fungi in the order Erysiphales (Figure \(\PageIndex{10}\)).
.jpeg?revision=1&size=bestfit&width=415&height=312)
.jpeg?revision=1&size=bestfit&width=415&height=331)
Lichenized Fungi
- Explain the lichen symbiosis.
- Identify structures in the lichen thallus.
Lichens are fungi that live in a symbiotic association with a green alga or cyanobacterium (the "photobiont"), or both. The primary fungal partner (the "mycobiont") in most lichens (98% of them) is an ascomycete. Basidiomycetes make up the remainder. The relationship is often characterized as mutualistic; that is, both partners benefit. However, evidence (see "The British Soldier" below) suggests that while the fungus is dependent on its autotrophic partner, the photobiont is often perfectly content to live alone. Recently many lichens have been found to harbor a second fungal partner, a basidiomycete yeast. Its function remains to be discovered, though the presence of the basidiomycete can change the outward appearance of the lichen (e.g. color).
The British Solder
The below image is of the colorful lichen called British soldier. The fungus is Cladonia cristatella, an ascomycete. Its name is the name given to the lichen. The photobiont is Trebouxia erici, a green alga. It is found in many other lichens as well, and also can be found growing independently. The algal cells eventually are killed by the fungus, but are continuously replaced by new ones. So, the relationship in this lichen is one of controlled parasitism rather than mutualism.
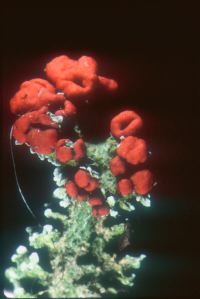
The red cap produces the spores of the fungus, but these alone cannot form new lichens. Other structures (e.g., soredia), containing both partners, are needed to disperse the lichen to new locations. Some lichens release only fungal spores. These mycobionts depend for their continued survival on finding an acceptable photobiont released from other lichens. Phylogenetic trees, based on both ribosomal RNA genes and many protein-coding genes, as well as fossils indicate that lichens have been present on the earth for at least 600 million years.
Today about 14,000 species of fungi are known to form lichens. Lichens display a range of colors and textures and can survive in the most unusual and hostile habitats, though they are extremely sensitive to air pollution.. They cover rocks, gravestones, tree bark, and the ground in the tundra where plant roots cannot penetrate. Lichens can survive extended periods of drought, become completely desiccated, and then rapidly become active once water is available again. In part because of this ability, as well as the pigments in some lichens that protect from UV radiation, lichens are some of the only living things to survive exposure to conditions in space.

Explore the world of lichens using this site from Oregon State University.

The body of a lichen, referred to as a thallus (Figure \(\PageIndex{13}\)), is formed of hyphae wrapped around the photosynthetic partner. The photosynthetic organism provides carbon and energy in the form of carbohydrates. If cyanobacteria are involved, they fix nitrogen from the atmosphere, contributing nitrogenous compounds to the association. In return, the fungus supplies minerals and protection from dryness and excessive light by encasing the algae in its mycelium. The fungus also attaches the symbiotic organism to the substrate.
The thallus of lichens grows very slowly, expanding its diameter a few millimeters per year. Both the fungus and the alga participate in the formation of dispersal units for reproduction. Some lichens produce soredia (Figure \(\PageIndex{14}\)), clusters of algal cells surrounded by mycelia, for asexual reproduction. Soredia are dispersed by wind and water and form new lichens. To reproduce sexually, the fungus makes spores that must find a new photosynthetic partner shortly after germinating.

Basidiomycota
- Use life history traits and morphological features to distinguish between Ascomycota and Basidiomycota
- Identify the parts of a mushroom
- Identify structures in the Basidiomycota life cycle and know their ploidy
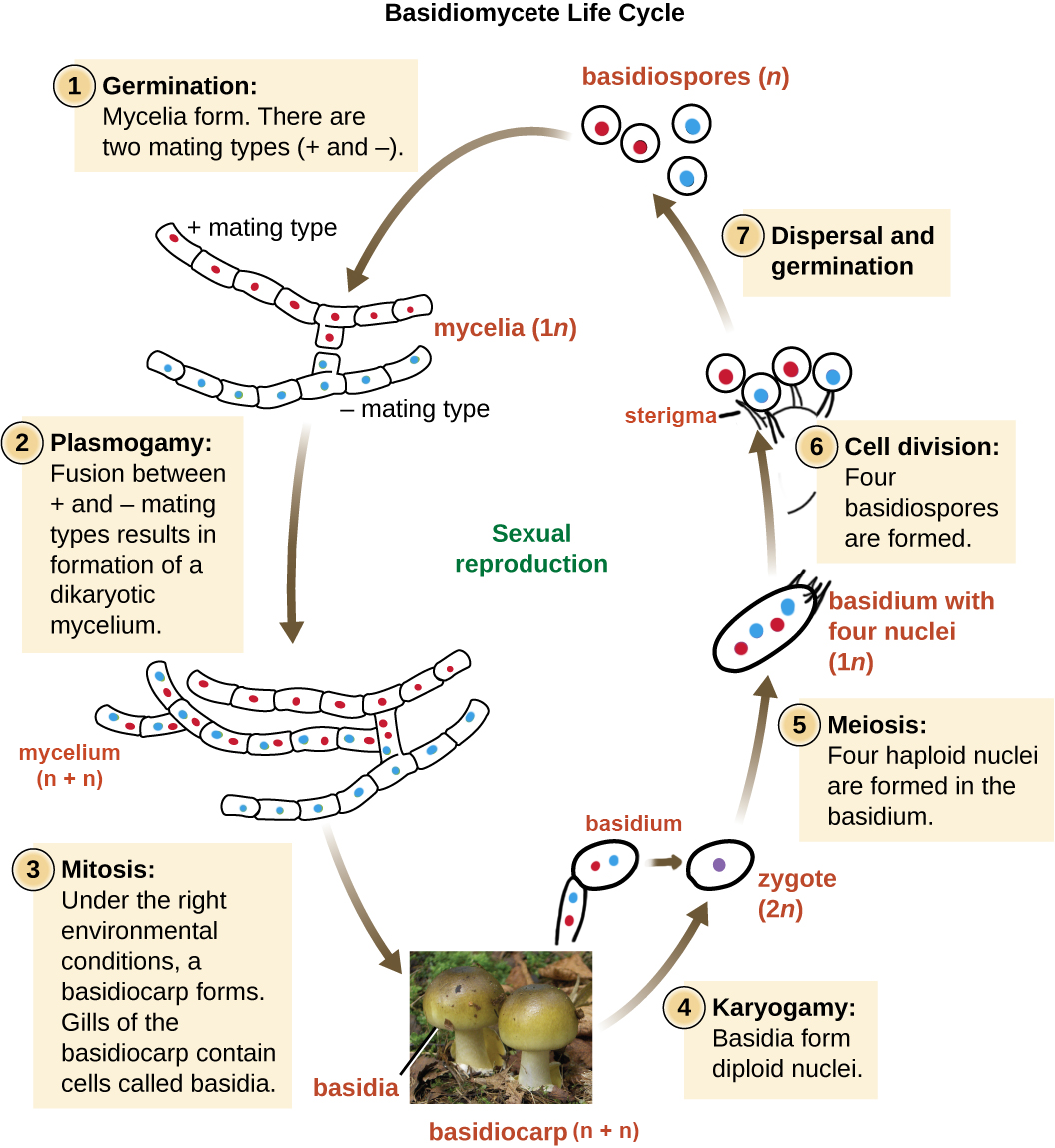
The Basidiomycota (basidiomycetes) are fungi that produce haploid basidiospores (spores produced through budding) from club-shaped cells called basidia (Figure \(\PageIndex{13}\)). These are typically formed within fruiting bodies called basidiocarps. They are important as decomposers (particularly of wood), plant pathogens, mutualists, and food sources for many animals. Basidiomycetes include the group of fungi that forms mushrooms, the rusts, and the smuts. Mushrooms are basidiocarps formed from masses of interwoven hyphae growing up from the mycelium. The basidia develop on fertile surfaces of the mushroom and release their spores (typically four from each basidium) into the air.
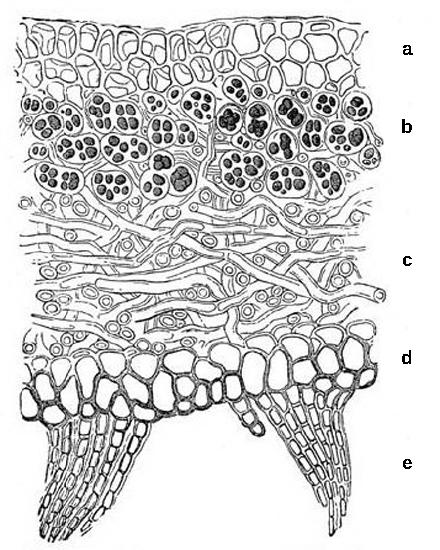
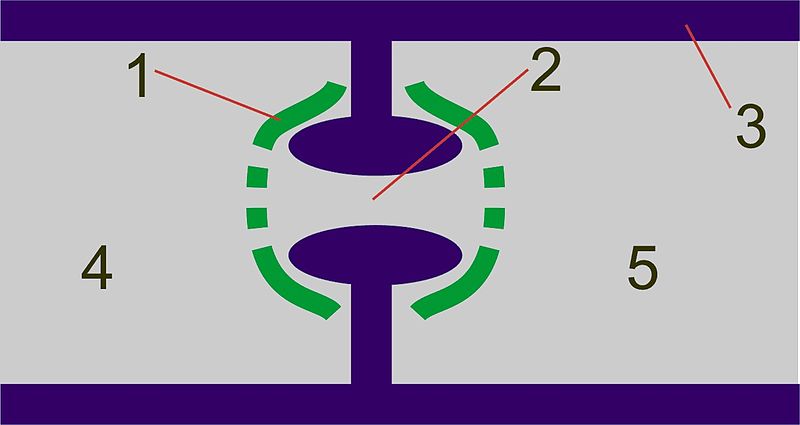
They hyphae of basidiomycetes are septate with clamp connections where the septa form (Figure \(\PageIndex{14}\)). The septal structure is more complex than in ascomycetes. The septum is swollen around the pore (a dolipore septum) and is flanked by structures called parenthesomes (Figure \(\PageIndex{15}\)).
Fungi with the following structures can be placed in the Basidiomycota*:
*It is important to note that these features may look different or not be present at all in some groups of Basidiomycota, such as the rusts (Pucciniomycotina) and smuts (Ustilagomycotina) -- see Chapter 3.6.4: Rusts & Smuts in the Photographic Atlas. However, there are many physiological and genetic similarities that support grouping these organisms together in the Basidiomycota.
Basidia and Basidiospores
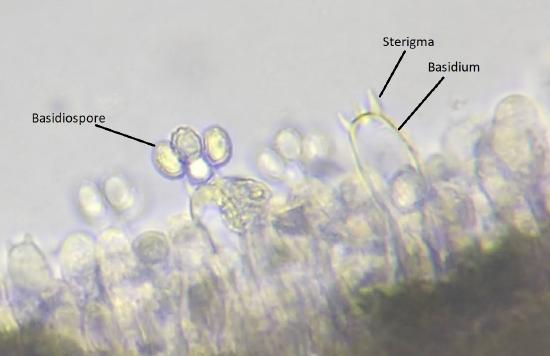
Both karyogamy and meiosis occur within a cell called the basidium (Figure \(\PageIndex{15}\)). Haploid basidiospores from atop projections on the basidum called sterigmata (sing. sterigma). There are generally four spores, as shown in the image below, though the number of spores produced can vary by species. For example, the mushroom you are likely most familiar with, Agaricus bisporus (though you probably know it as a crimini or button mushroom at its immature stage and portobello at maturity), only produces two spores on each basidium (bi- meaning two).
Clamp Connections
Basidiomycetes maintain their dikaryotic (n+n) state in each hyphal compartment by making structures called clamp connections (Figure \(\PageIndex{16}\)). These are not always present, but provide a helpful identification feature when they are!
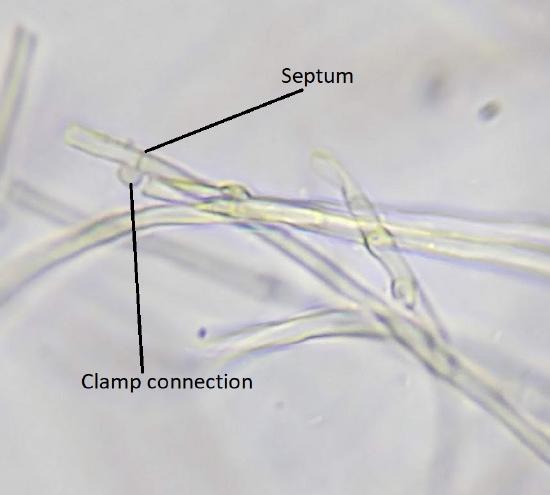
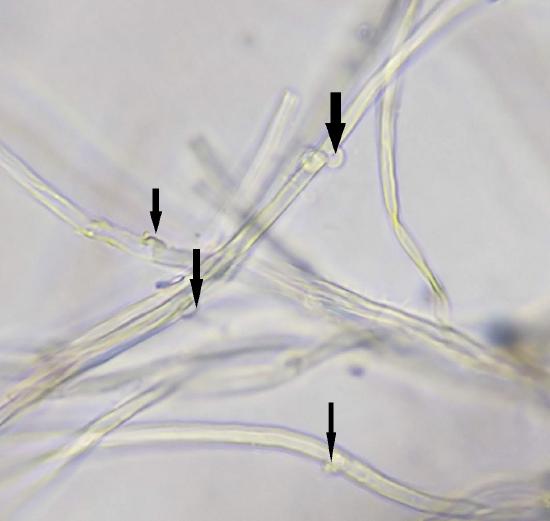
Complex Septations
Complex septations are shown in Figure \(\PageIndex{17}\).
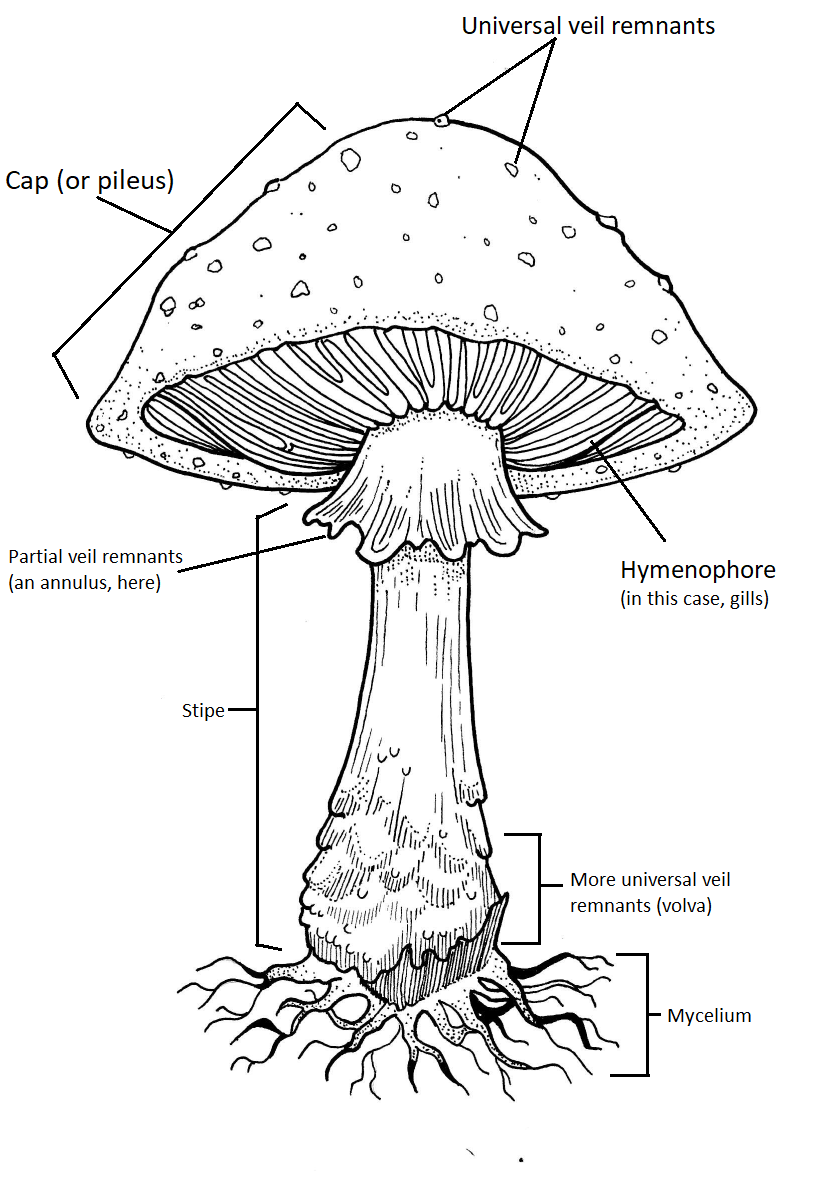
General Mushroom Anatomy
Though only a subset of basidiocarps look this way, they are the model for how we describe "mushrooms". In mycology, this type of basidiocarp is called "agaricoid" or "agaric" because it is the general form we see in the genus Agaricus. A more complex version of the agaric mushroom is seen in the genus Amanita (Figure \(\PageIndex{18}\)).
Basidiomycete Life Cycle
The life cycle of basidiomycetes involves an extension of the dikaryotic phase (Figure \(\PageIndex{19}\)). Mycelia of different mating strains combine (plasmogamy) soon after germination and produce a secondary, dikaryotic mycelium that contains haploid nuclei of two different mating strains (a dikaryon). This is the dikaryotic stage of the basidiomycete life cycle, and it is the dominant stage. Eventually, the secondary mycelium generates a basidiocarp. The basidiocarp can vary greatly in morphology, but in the sense of a standard mushroom (Figure \(\PageIndex{15}\)), the developing basidia are produced on the surface of gills located under cap.Within the club-shaped basidium, meiosis occurs and a diploid zygote is formed (karyogamy). The zygote divides by meiosis to produce four haploid nuclei. The haploid nuclei migrate into basidiospores, which germinate and generate monokaryotic hyphae. The mycelium that results is called a primary mycelium.
Summary
Macrofungi are represented by two major groups that can form macroscopic fruiting structures. However, many lineages within these groups might only reproduce asexually, form yeasts, or form microscopic fruiting structures (such as the rusts and smuts).
Ascomycetes typically form 8 ascospores within a structure called an ascus. In most lineages, these are produced within an ascocarp, which can be an apothecium (cup), perithecium (flask), or cleistothecium (ball). Ascomycetes have hyphae with simple septations. Their life cycle involves an extended dikaryotic phase that takes place within the ascocarp, forming dikaryotic ascogenous hyphae. These hyphae will eventually form asci, where karyogamy will take place, followed shortly after by meiosis, and usually mitosis.
Basidiomycetes typically form 4 basidiospores externally on a basidium. In the Agaricomycotina, these are produced on or in a basidiocarp, what we call mushrooms, specialized for spore dispersal. The basidiomycete life cycle is almost entirely dikaryotic. Haploid spores germinate, form a monokaryon, then must fuse with another monokaryon shortly afterward. These forms a dikaryotic mycelium with dolipore septations and clamp connections. Karyogamy only takes place within the basidia, follow promptly by meisosis to produce basidiospores.
Both ascomycetes and basidiomycetes for relationships with photosynthetic partners, such as algae or cyanobacteria, to form lichens. In this mutualism, the photobiont provides sugars from photosynthesis, while the mycobiont forms a protective thallus. In the case of cyanobacteria, these may also provide nitrogen fixation.
Attributions
Curated and authored by Maria Morrow, CC-BY-NC, using the following sources:
- 19.1.7 Fungi from Biology by John. W. Kimball (licensed CC-BY)
- 24.2 Classifications of Fungi and 24.3 Ecology of Fungi from Biology 2e by OpenStax (licensed CC-BY). Access for free at openstax.org.
- 5.3 Fungi from Microbiology by OpenStax (licensed CC-BY). Access for free at openstax.org.