
18.5: The Role of Consumers and Alternative Stable States
- Page ID
- 81423
18.5.1: Succession in Consumers
Animal communities also undergo change throughout successional time. In the lichen stage fauna is sparse. It comprises a few mites, ants and spiders living in cracks and crevices. The fauna undergoes a qualitative increase during the herb grass stage. The animals found during this stage include nematodes, insects larvae, ants, spiders, mites, etc. The animal population increases and diversifies with the development of the forest climax community. The fauna consists of invertebrates like slugs, snails, worms, millipedes, centipedes, ants, bugs; and vertebrates such as squirrels, foxes, mice, moles, snakes, various birds, salamanders and frogs. The pioneering fauna will colonize an area only after flora and fungi have inhabited the area. Soil fauna, ranging from microscopic protists to larger invertebrates, have a role in soil formation and nutrient cycling. Bacteria and fungi are the most important groups in the breakdown of organic detritus left by primary producing plants such as skeletal soil, moss and algae. Soil invertebrates enhance fungal activity by breaking down detritus. As soil develops, earthworms and ants alter soil characteristics. Worm burrows aerate soil and ant hills alter sediment particle size dispersal, altering soil character profoundly.
Though vertebrates in general would not be considered pioneer species, there are exceptions. Natterjack toads are specialists in open, sparsely vegetated habitats which may be at an early seral stage.1 Wide-ranging generalists visit early succession stage habitats, but are not obligate species of those habitats because they use a mosaic of different habitats. Vertebrates can effect early seral stages. Herbivores may alter plant growth. Fossorial mammals could alter soil and plant community development. In a profound example, a seabird colony transfers considerable nitrogen into infertile soils, thereby altering plant growth. A keystone species may facilitate the introduction of pioneer species by creating new niches. For example, beavers may flood an area, allowing new species to immigrate.2
Succession of micro-organisms including fungi and bacteria occurring within a microhabitat is known as microsuccession or serule. Like in plants, microbial succession can occur in newly available habitats (primary succession) such as surfaces of plant leaves, recently exposed rock surfaces (i.e., glacial till) or animal infant guts,3 and also on disturbed communities (secondary succession) like those growing in recently dead trees, decaying fruits,4 or animal droppings.
18.5.2: Alternative Stable States
One of the main ways consumers can affect succession is through the redirection of successional trajectories. By preferentially consuming some species, consumers can change the direction of succession, resulting in a different community type than would typically be predicted. Expected successional trajectories are predicted based on the observation of community changes through time, or through space-for-time substitutions where stands of different ages are compared, with the expectation that the youngest stands will grow to look like the oldest as time passes. However, given substantial perturbations, communities may end up going down an entirely different path than we would predict. Sometimes, they will even get "stuck" in this state, and never return to look as we would expect. This becomes especially problematic when they place they get "stuck" is low-diversity or dominated by invasive species, and makes it very hard to restore healthy ecosystems.
We can visualize a community about to undergo succession as a ball about to roll down a hill. The trajectory of succession is predictable, and gravity will pull the ball to the bottom of the hill. Disturbance can provide energy to push the ball back up the hill, but it should consistently return to the same stable end point. However, the ball may exist in a landscape with many low points, where the ball would have to roll far up a hill to change positions. Thus, there may be several alternative stable states where the community will be resistant to change. Catalysts, such as invasive species or ecosystem engineering animals can push the ball over a peak and into a different valley, resulting in a different endpoint to our successional trajectory. At this point, even a disturbance may not provide enough energy to push the ball all the way back up the hill and over a peak into the original successional trajectory. The new endpoint is stable, and hard to restore from.
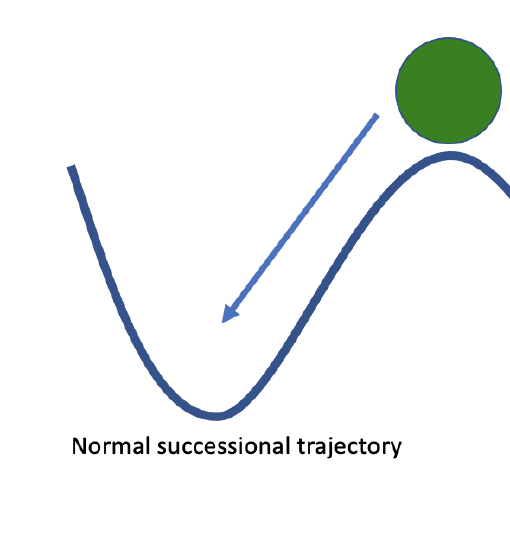
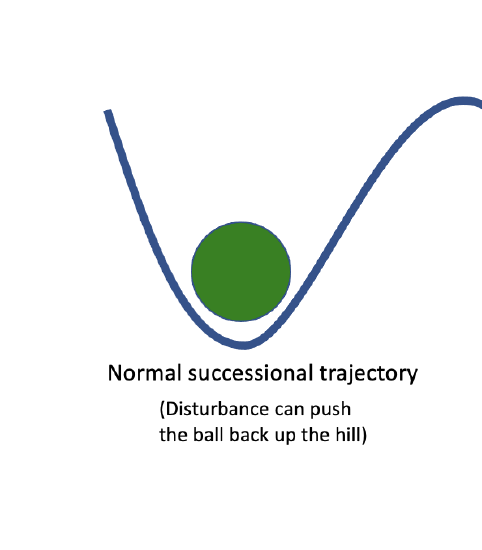
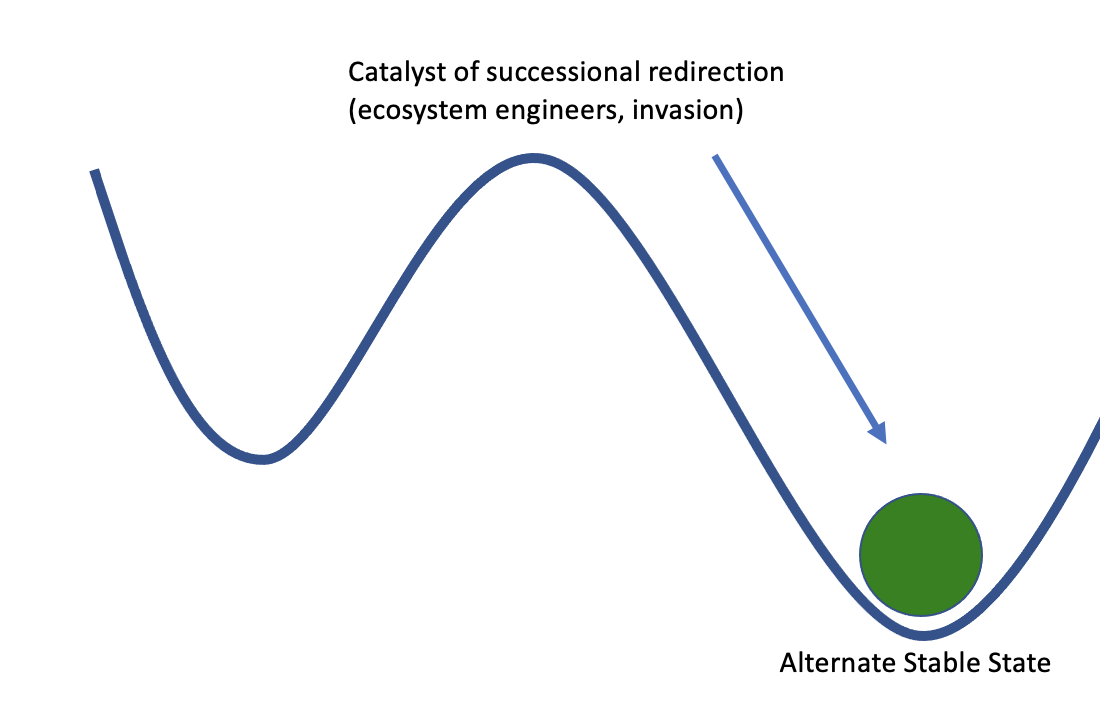
Figure \(\PageIndex{1}\): a) the starting point of early succession, which should proceed "downhill" along a predictable trajectory. b) the stable climax community at the end of a successional trajectory. c) some catalyst has pushed our community out of its original state and into an alternative stable community state.
As the cover of forested land increases and the climate warms, moose (Alces alces americana) are expanding their range on the northern and southern edges, even as far south as Connecticut5. Moose often spend time in early-successional forests because of the high-availability of young tree and shrub growth to browse on. Moose not only browse the tips of trees, reducing their ability to grow vertically, but they can actually break trees in half as they try to reach the palatable tips of branches and leaves. As a result, moose may be redirecting the trajectory of early-successional forests, especially in the southernmost edges of their range, in Connecticut, by preferentially browsing certain species of trees while avoiding others6.
A typical successional trajectory for a mixed-hardwood forest in Connecticut would look something like this:

Figure \(\PageIndex{2A}\): Typical successional trajectory.
However, when moose browsing is added to the equation, we would expect one of two possible outcomes.
1. If moose browse all species equally and they are sufficiently abundant, they may actually delay succession, keeping the forest in a more-open, shorter state (the ball takes longer to roll down the hill).

Figure \(\PageIndex{2B}\): Possible outcome of typical successional trajectory when moose are added to the scenario, where the moose delay the succession development rate.
2. If moose browse preferentially, avoiding species they dislike (such as Spruce), they may redirect the trajectory of the forest. The mature forest which eventually takes hold will be dominated by these browse-resistant tree species, and the community composition will look very different than the trajectory in the absence of moose.

Figure \(\PageIndex{2C}\): Alternate possible outcome of typical successional trajectory when moose are added to the scenario, where the moose redirect the species diversity.
Both of these outcomes occur where moose are present, but which outcome we see is contingent on the abundances of preferred and non-preferred tree species. In the first case, if moose are controlled or removed in the third time point, the closed forest should eventually look like the original successional trajectory. However, in the second case, even if moose are no longer present in the third time point (this is likely, because moose frequent open, younger forests), the forest will still be dominated by browse-tolerant unpalatable tree species. It will no longer resemble the expected successional trajectory, and it will be unlikely to revert without major disturbance. The ability of moose to redirect succession in this way makes them an example of an ecosystem engineer. In the second case, by keeping forests open and in an early successional stage for longer, moose provide habitat and maintain food resources for many species of bird and other small animals.
References
- Faucher, L., Hénocq, L., Vanappelghem, C., Roundel, S., Tocqueville, R., Galina, S., Godé, C., Jaquiéry, J., & Arnaud, J.F. (2017). When new human-modified habitats favor the expansion of an amphibian pioneer species: Evolutionary history of the natterjack toad (Bubo calamity) in a coal basin. Molecular Ecology, 26(17), pp. 4434–4451. doi:10.1111/mec.14229. ISSN 1365-294X. PMID 28667796. S2CID 25656968.[permanent dead link]
- Wallwork, J.A. (1970). Ecology of soil animals. McGowan-Hill. ISBN 978-0070941250.
- Ortiz-Álvarez, R., Fierer, N., de los Ríos, A., Casamayor, E.O., Barberán, A. (2018). Consistent changes in the taxonomic structure and functional attributes of bacterial communities during primary succession. The ISME Journal, 12(7), pp. 1658–1667. doi:10.1038/s41396-018-0076-2. ISSN 1751-7370. PMC 6018800. PMID 29463893.
- Martin, P.L., King, W., Bell, T.H., & Peter, K. (2021). The decay and fungal succession of apples with bitter rot across a vegetation diversity gradient. Phytobiomes Journal: PBIOMES–06–21-0039-R. doi:10.1094/PBIOMES-06-21-0039-R. ISSN 2471-2906.
- http://wildlifeofct.com/moose.html
- Olmsted, C.F., Betras, T.L., Pasquini, S.C., DeStephano, S., Faison, E.K., & Carson, W.P. (2021). Characteristics of stem-breaking by moose (Alces alces, Cervidae): A case-study and worldwide review. The Journal of the Torrey Botanical Society, 147(4), pp. 304-315.
Contributors and Attributions
Modified by Castilleja Olmsted (University of Pittsburgh) and Kyle Whittinghill (University of Vermont) from the following sources:
- Original text for Alternative Stable States and Box 1, ball diagrams, by Castilleja Olmsted.
- Forest illustrations for Box 1 by Tiffany Betras, reproduced and modified here with permission.
- https://en.wikipedia.org/wiki/Ecological_succession
- https://en.wikipedia.org/wiki/Pioneer_species