
17.3: Evolution of Mutualisms
- Page ID
- 80944
Evolution of Mutualisms
Many mutualisms are thought to have evolved from antagonistic interactions. Several studies have determined ecological or evolutionary conditions driving transitions from mutualism to antagonism (mutualism breakdown) (Kiers et al. 2010; Sachs and simms 200), yet limited theory has been developed to mechanistically explain the evolutionary transitions from antagonism to mutualism (but see de Mazancourt et al. 2001; deMazancourt et al. 2005). Models of virulence predict evolution of reduced antagonism, perhaps toward mutualism, in cases of parasitism where host fidelity is high and availability of alternate hosts is low (Yamamura 1993; Yamamura 1996). In these models, host fidelity is achieved when lineages of parasites and their host are tightly linked through vertical transmission from parent to offspring.
Compelling evidence for the ‘virulence theory’ comes from interactions involving microbial parasites (Weeks et al. 2007; Sachs et al. 2014). For example, Wolbachia are endosymbiotic bacteria that live inside the cells of their invertebrate hosts. They are transmitted directly from mother to offspring, and spread through populations by manipulating the reproduction of their hosts. The most common reproductive manipulation responsible for the spread of these bacteria, called “cytoplasmic incompatibility,” arises when infected males mate with uninfected females, resulting in fewer offspring than normal. There are fitness costs for the hosts associated with Wolbachia infections, most commonly involving a reduction in egg production. The virulence theory predicts and evidence suggests that there is selection for the bacteria to evolve a more benign lifestyle, changing the bacterium from being parasitic to more mutualistic, where Wolbachia-infected hosts have higher rates of egg production than their uninfected counterparts (Weeks et al. 2007).
Reciprocal evolutionary responses are most likely important to the evolution of symbiotic mutualisms, and subsequent diversification may be the result of the mutualism improving niche width, population size, or transition into adaptive zones (Hembry et al 2014). However, with the exception of plant-pollinator interactions, it is unclear how important mutualistic coevolution is as a driving force of diversification, especially as compared to the well-established mechanisms associated with competitive and antagonistic coevolution.
Plant-pollinator interactions are powerful tools of evolution because pollinators transport gametes (via pollen) between flowers. Plant-pollinator relationships are thought to be critical to the historical diversification of angiosperm flowers. In fact, up to 25% of plant speciation events could be as a result of interactions with pollinators (van der Niet and Johnson 2012).
The obligate pollinating seed-predation mutualisms of the yucca and yucca moth and fig and fig wasps are attributed to coevolution diversification because the intimacy of the association may prevent species from making phylogenetically drastic switches to new partners. Interacting with a distant relative of your partner may not be an option if coevolution has created incompatibilities in specialized traits required for the interaction (Figure \(\PageIndex{1}\)). For instance, if laying eggs into yucca flowers requires a particular complement of ovipositor traits and moth behaviors, we might predict to find host switches only between closely related yuccas. Initial analyses of yucca and yucca moth phylogenies suggest that the trees are somewhat congruent, although there have been at least two instances of moths switching hosts to quite distantly related yuccas (Pellmyr 2003). The interaction between yuccas and yucca moths is clearly one that is highly specialized and coevolved, yet we do not find a phylogenetic pattern of perfect matching between plant and insect.
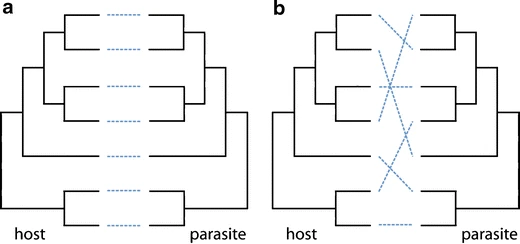
Figure \(\PageIndex{1}\): Comparison of host and parasite phylogenetic trees. The dotted lines indicate host associations of the parasites. a) Matching speciation events between host and parasite show a strict pattern of cospeciation. b) Host switches to distant relatives are shown by the crossing lines. Some parasites use closely related hosts, but most have jumped to a distant relative. Figure from Segraves 2010.
However, there is also evidence opposing the primacy of mutualistic coevolution in these relationships. Figs have been shown to be successfully pollinated regardless of pollinator species (Moe and Weiblen 2012) and pollinator host-switch may be more frequent than co-speciation events (Wang et al. 2021; Figure \(\PageIndex{2}\)). There is also evidence that geographic isolation plays a significant role in yucca diversification (Althoff et al. 2012; Smith et al. 2008). These together suggest that while coevolution plays a role in the emergence of these mutualisms, it is not the exclusive mechanism and may not even be the primary one.
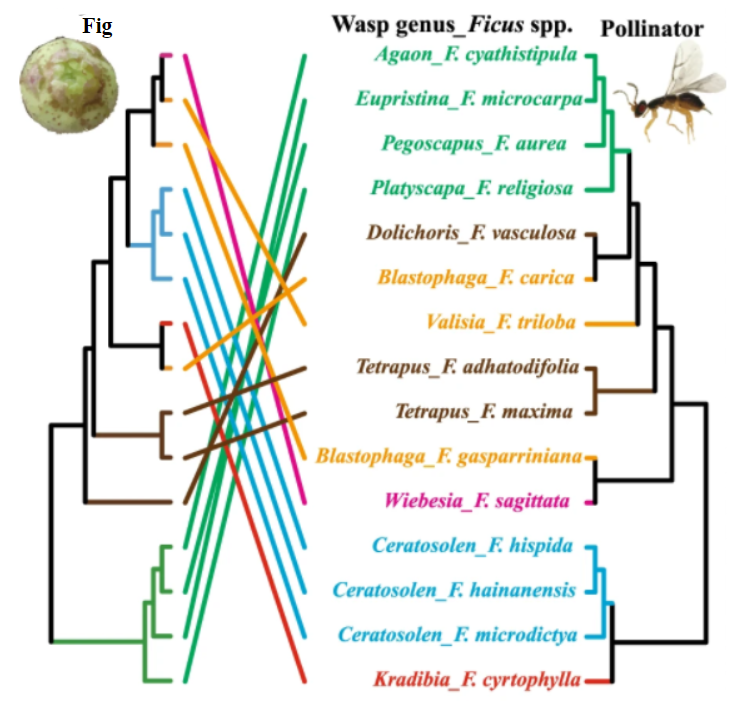
Figure \(\PageIndex{2}\): Cophylogenetic comparison of the pollinators associated with the 15 fig species studied. Ficus and pollinator phylogenies used were indicated with symbol of figs and wasp separately. Tip-name colors indicate subgeneric classification. Extensive phylogenetic incongruence is shown for each pair of phylogenies. Figure modified from Wang et al. 2021.
References
Althoff, D.M., Segraves, K.A., Smith, C.I., Leebens-Mack, J., & Pellmyr, O. (2012). Geographic isolation trumps coevolution as a driver of yucca and yucca moth diversification. Molecular Phylogenetics and Evolution, 62, pp. 898–906.
de Mazancourt, C., Loreau, M., & Dieckmann, U. (2001). Can the evolution of plant defense lead to plant‐herbivore mutualism? Am. Nat., 158, pp. 09–123.
de Mazancourt, C., Loreau, M., & Dieckmann, U. (2005). Understanding mutualism when there is adaptation to the partner. J. Ecol., 93, pp. 305–314.
Hembry, D.H., Yoder, J.B., & Goodman, K.R. (2014). Coevolution and the diversification of Llfe. The American Naturalist, 184(4), pp. 425–38. https://doi.org/10.1086/677928.
Kiers, T.E., Palmer, T.M., Ives, A.R., Bruno, J.F. & Bronstein, J.L. (2010). Mutualisms in a changing world: An evolutionary perspective. Ecol. Lett., 13, pp. 1459–1474.
Moe, A.M., & Weiblen., G.D. (2012). Pollinator-mediated reproductive isolation among dioecious fig species (Ficus, Moraceae). Evolution, 66, pp. 3710–3721.
Pellmyr, O. (2003). Yuccas, yucca moths, and coevolution: A review. Ann. Mo. Bot. Gard., 90, pp. 35–55.
Sachs, J. & Simms, E. (2006). Pathways to mutualism breakdown. Trends Ecol. Evol., 21, pp. 585–592.
Sachs, J., Skophammer, R., Bansal, N. & Stajich, J. (2014). Evolutionary origins and diversification of proteobacterial mutualists. Proc. R. Soc. B Biol. Sci., 281, 20132146.
Smith, C.I., Pellmyr, O., Althoff, D.M., Balcázar-Lara, M., Leebens-Mack, K., & Segraves, K.A. (2008). Pattern and timing of diversification in Yucca (Agavaceae): Specialized pollination does not elevate rates of diversification. Proceedings of the Royal Society B: Biological Sciences, 275, pp. 249–258.
van der Niet, T., & Johnson, S.D. (2012). Phylogenetic evidence for pollinator-driven diversification of angiosperms. Trends in Ecology and Evolution, 27, pp. 353–361.
Weeks, A., Turelli, M., Harcombe, W., Reynolds, K. & Hoffmann, A. (2007). From parasite to mutualist: Rapid evolution of Wolbachia in natural populations of Drosophila. PLoS Biol., 5, e114.
Yamamura, N. (1993). Vertical transmission and evolution of mutualism from parasitism. Theor. Popul. Biol., 44, pp. 95–109.
Yamamura, N. (1996). Evolution of mutualistic symbiosis: A differential equation model. Res. Popul. Ecol., 38, pp. 211–218.
Contributors and Attributions
This chapter was written by Aaron Howard with text taken from the following CC-BY resources:
-
Johnson, C.A., Smith, G.P., Yule, K. et al. (2021). Coevolutionary transitions from antagonism to mutualism explained by the Co-Opted Antagonist Hypothesis. Nat Commun, 12, 2867. https://doi.org/10.1038/s41467-021-23177-x
-
Segraves, K.A. (2010). Branching out with coevolutionary trees. Evo. Edu. Outreach, 3, pp. 62–70. https://doi.org/10.1007/s12052-009-0199-z
-
Wang, G., Zhang, X., Herre, E.A. et al. (2021). Genomic evidence of prevalent hybridization throughout the evolutionary history of the fig-wasp pollination mutualism. Nat Commun., 12, 718. https://doi.org/10.1038/s41467-021-20957-3
-
Weeks, A., Turelli, M., Harcombe, W., Reynolds, K. & Hoffmann, A. (2007). From parasite to mutualist: Rapid evolution of Wolbachia in natural populations of Drosophila. PLoS Biol., 5, e114.