
9.4: Disturbance
- Page ID
- 103380
Ecological Disturbance
Ecological disturbance, is a temporary change in environmental conditions that causes a pronounced change in a community. Disturbances often act quickly and with great effect, to alter the physical structure or arrangement of biotic and abiotic elements. Disturbance plays a significant role in shaping the structure of individual populations and the biodiversity of whole ecosystems.
Minor disturbances include localized wind events, droughts, floods, small wildland fires, and disease outbreaks in plant and animal populations. In contrast, major disturbances include large-scale wind events (such as tropical cyclones), volcanic eruptions, tsunamis, intense forest fires, epidemics, ocean temperature changes stemming from El Niño events or other climate phenomena, and the devastating effects of human impact on the environment (anthropogenic disturbances) such as clear cutting, pollution, or the introduction of invasive species. Not only invasive species can have a profound effect on an ecosystem, but also naturally occurring species can cause disturbance by their behavior.
Disturbance forces can have profound immediate effects on ecosystems and can, accordingly, greatly alter the natural community’s population size or species richness. Because of these and the impacts on populations, disturbance determines the future shifts in dominance, various species successively becoming dominant as their life history characteristics, and associated life-forms, are exhibited over time. The notion of ecological disturbance has deep historical roots in ecological thinking; the first conceptual disturbance-related model in modern ecology was ecological succession, an idea emphasizing the progressive changes in ecosystem structure that follow a disturbance.



Impact
The ecological impact of a disturbance is dependent on its intensity and frequency, on the spatial distribution (or the spatial pattern) and size of the disturbed patches, and on the scale (the spatial extent) of the disturbance. Some disturbances may by cyclic. Fire disturbances, for example, occur more often in areas with a higher incidence of lightning and flammable biomass. Other disturbances, such as those caused by humans, invasive species or impact events, can occur anywhere and are not necessarily cyclic. When multiple disturbance events affect the same location in quick succession, this often results in a "compound disturbance," an event which, due to the combination of forces, creates a new situation which is more than the sum of its parts. For example, windstorms followed by fire can create fire temperatures and durations that are not expected in even severe wildfires, and may have surprising effects on post-fire recovery. Diversity is generally low in harsh environments (with frequent or compound disturbances) because of the intolerance of all but opportunistic and highly resistant species to such conditions.
The spatial scale of natural disturbances, which is known to span about 10 orders of magnitude, is also important. For example, a drought that might devastate microbes in a temporary pond would be inconsequential to an elephant. A single tree uprooted by a hurricane is a disaster for the resident ants, but it may become a necessary resource for forest frogs as sufficient water collects around the root cavity. Likewise, while a fire may decimate wildlife populations and scorch large areas of land, the cones of the northern jack pine (Pinus banksiana), which are tiny in comparison, require the presence of fire to open (Figure \(\PageIndex{1}\)).
Recovery
The recovery process for species removed by a disturbance is critically dependent on its dispersal capability and the distance between the disturbed site and surviving source populations. Immediately after a disturbance there is a pulse of recruitment or regrowth under conditions of little competition for space or other resources. These conditions may favor the success of different species than the pre-disturbed community. After the initial pulse, recruitment slows since once an individual species is established it may be very difficult to displace. However, in the absence of further disturbance forces, many communities trend back toward pre-disturbance conditions. Long lived species and those that can regenerate in the presence of their own adults finally become dominant.
Some species are particularly suited for exploiting recently disturbed sites. Vegetation with the potential for rapid growth can quickly take advantage of the lack of competition. In the northeastern United States, shade-intolerant trees like pin cherry and aspen quickly fill in forest gaps created by fire or windstorm (or human disturbance). Silver maple and eastern sycamore are similarly well adapted to floodplains. They are highly tolerant of standing water and will frequently dominate floodplains where other species are periodically wiped out. Species that are well adapted for exploiting disturbance sites are referred to as pioneers. They usually grow and reproduce quickly, but may not be the strongest competitors and are usually replaced by other, more competitively dominant species over time.
The interplay between disturbance and biological processes seems to account for a major portion of the organization and spatial patterning of natural communities. Although disturbances tend to negatively affect populations of resident plants, animals, and other organisms in a given ecosystem, they provide some fugitive species with opportunities to move into and gain footholds in ecosystems whose biological communities once excluded them. This process results in an increase in the biodiversity of the ecosystem.
Intermediate Disturbance Hypothesis
Disturbance variability and species diversity are heavily linked. The success of a wide range of species from all taxonomic groups is closely tied to natural disturbance events such as fire, flooding, and windstorm. Diversity tends to be low in harsh environments because of the intolerance of all but opportunistic and highly resistant species to such conditions. The interplay between disturbance and these biological processes seems to account for a major portion of the organization and spatial patterning of natural communities.
The intermediate disturbance hypothesis (IDH) suggests that local species diversity is maximized when ecological disturbance is neither too rare nor too frequent. At low levels of disturbance, more competitive organisms will push subordinate species to extinction and dominate the ecosystem. At high levels of disturbance, due to frequent forest fires or human impacts like deforestation, all species are at risk of going extinct. According to IDH theory, at intermediate levels of disturbance, diversity is thus maximized because species that thrive at both early and late successional stages can coexist (Figure \(\PageIndex{2}\)). IDH is a nonequilibrium model used to describe the relationship between disturbance and species diversity.
IDH is based on the following premises:
- ecological disturbances have major effects on species richness within the area of disturbance,
- interspecific competition results from one species driving a competitor to extinction and becoming dominant in the ecosystem, and
- moderate ecological scale disturbances prevent interspecific competition.
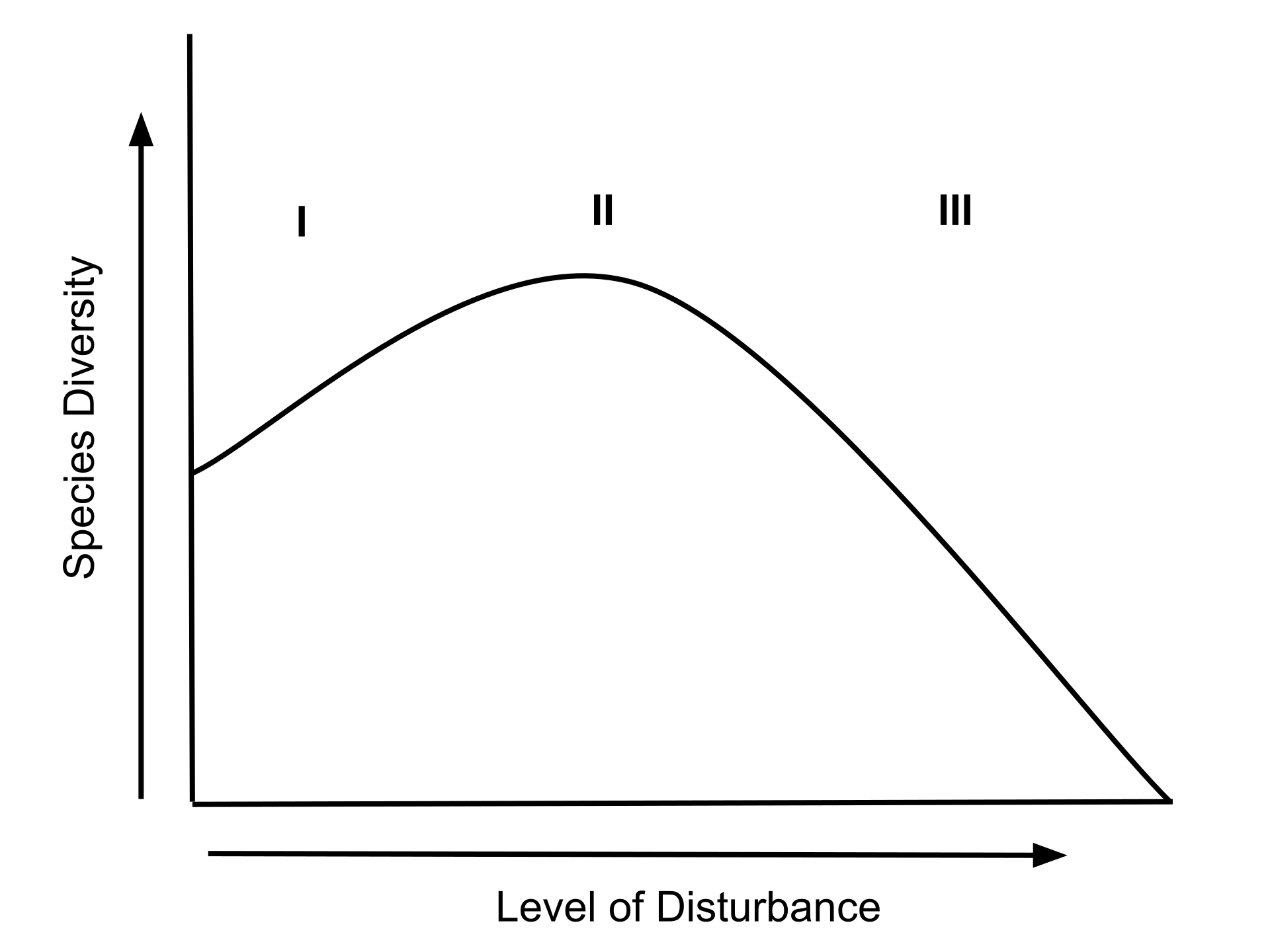
Disturbances act to disrupt stable ecosystems and clear species' habitat. As a result, disturbances lead to species movement into the newly cleared area. Once an area is cleared there is a progressive increase in species richness and competition between species takes place. Once the conditions that create a disturbance are gone, and competition between species in the formerly disturbed area increases, species richness decreases as competitive exclusion increases. Another explanation is that proposed by Joseph H. Connell who suggested that relatively low disturbance leads to decreased diversity while high disturbance causes an increase in species movement. These proposed relationships lead to the hypothesis that intermediate disturbance levels would be the optimal amount of disorder within an ecosystem.
Another way of thinking about the IDH requires that we consider the types of organisms that could specialize in areas with different levels of disturbance. K-selected species generally demonstrate more competitive traits. Their primary investment of resources is directed towards growth, causing them to dominate stable ecosystems over a long period of time. In contrast, r-selected species colonize open areas quickly and can dominate landscapes that have been recently cleared by disturbance. These characteristics attribute to the species that thrive in habitats with higher and lower amounts of disturbance. Based on the contradictory characteristics of both of these examples, areas of occasional disturbance allow both r and K species to flourish in the same area. If K-selected and r-selected species can live in the same region, species richness can reach its maximum.
References
- Dial, R.; Roughgarden, J. (1988). "Theory of marine communities: the intermediate disturbance hypothesis". Ecology. 79 (4): 1412–1424.
- Wilkinson, David M. (1999). "The Disturbing History of Intermediate Disturbance". Oikos. 84 (1): 145–7.
- Kricher, John C. (2011). Tropical Ecology. New Jersey, Princeton: Princeton University Press.
- Catford, Jane A.; Daehler, Curtis C.; Murphy, Helen T.; Sheppard, Andy W.; Hardesty, Britta D.; Westcott, David A.; Rejmánek, Marcel; Bellingham, Peter J.; et al. (2012). "The intermediate disturbance hypothesis and plant invasions: Implications for species richness and management". Perspectives in Plant Ecology, Evolution and Systematics. 14 (3): 231–41.
- Vandermeer, John; Boucher, Douglas; Perfecto, Ivette; de la Cerda, Inigo Granzow (1996). "A Theory of Disturbance and Species Diversity: Evidence from Nicaragua After Hurricane Joan". Biotropica. 28 (4): 600–13.
- Connell, J. H. (1978). "Diversity in Tropical Rain Forests and Coral Reefs". Science. 199 (4335): 1302–10.
- Hall, A. R.; Miller, A. D.; Leggett, H. C.; Roxburgh, S. H.; Buckling, A.; Shea, K. (2012). "Diversity-disturbance relationships: Frequency and intensity interact". Biology Letters. 8 (5): 768–71.
Attribution
This page is a modified derivative of:
- Disturbance (ecology) via Wikipedia (CC-SA 4.0)
- 18.2: What are the Effects of Disturbance? from Ecology for All! by Joshua Halpernunder a CC BY-NC-SA license