
11.2: DNA As the Molecule Responsible for Heredity
- Page ID
- 160284

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)- Explain how scientists established the link between DNA and heredity
- Describe key experiments (Griffith, Avery-MacLeod-McCarty, Hershey-Chase) that demonstrated DNA's role in heredity
DNA as the Molecule Responsible for Heredity
By the beginning of the 20th century, a great deal of work had already been done on characterizing DNA and establishing the foundations of genetics, including attributing heredity to chromosomes found within the nucleus. Despite all of this research, it was not until well into the 20th century that these lines of research converged and scientists began to consider that DNA could be the genetic material that offspring inherited from their parents. DNA, containing only four different nucleotides, was thought to be structurally too simple to encode such complex genetic information. Instead, protein was thought to have the complexity required to serve as cellular genetic information because it is composed of 20 different amino acids that could be combined in a huge variety of combinations. Microbiologists played a pivotal role in the research that determined that DNA is the molecule responsible for heredity.
Griffith’s Transformation Experiments
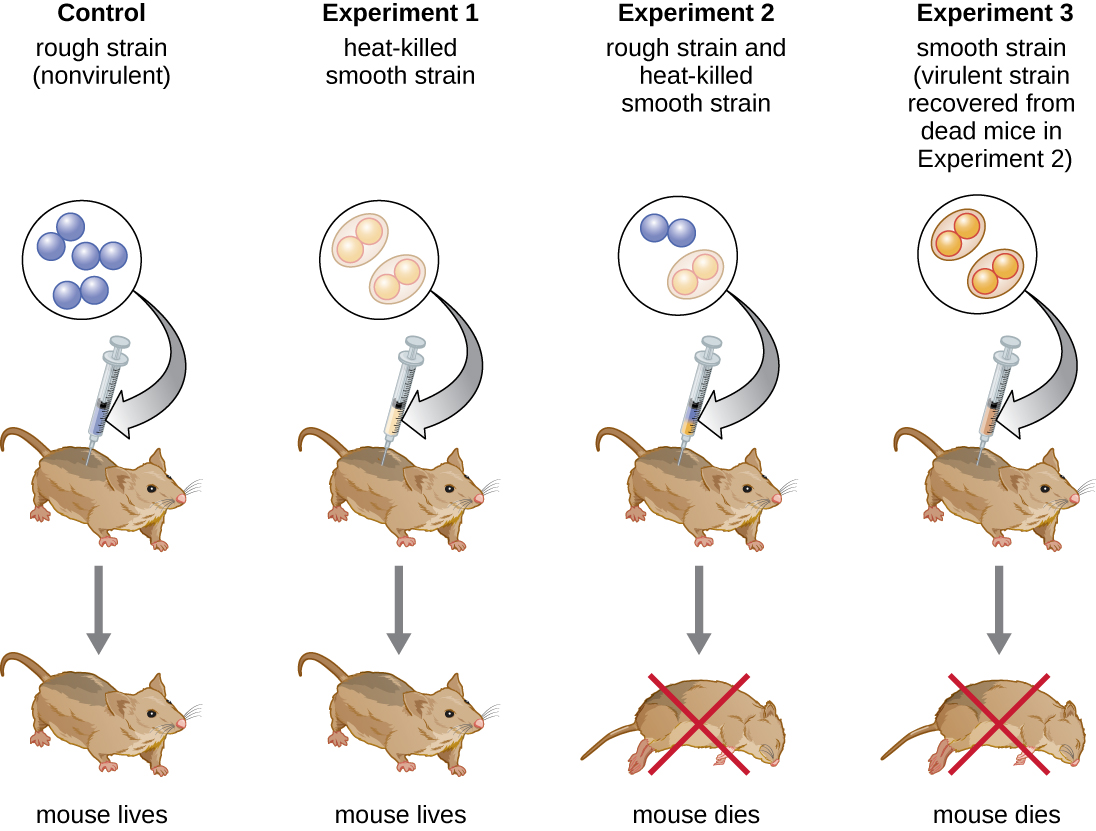
British bacteriologist Frederick Griffith (1879–1941) was perhaps the first person to show that hereditary information could be transferred from one cell to another “horizontally” (between members of the same generation), rather than “vertically” (from parent to offspring). In 1928, he reported the first demonstration of bacterial transformation, a process in which external DNA is taken up by a cell, thereby changing its characteristics.3 He was working with two strains of Streptococcus pneumoniae, a bacterium that causes pneumonia: a rough (R) strain and a smooth (S) strain. The R strain is nonpathogenic and lacks a capsule on its outer surface; as a result, colonies from the R strain appear rough when grown on plates. The S strain is pathogenic and has a capsule outside its cell wall, allowing it to escape phagocytosis by the host immune system. The capsules cause colonies from the S strain to appear smooth when grown on plates.
In a series of experiments, Griffith analyzed the effects of live R, live S, and heat-killed S strains of S. pneumoniae on live mice (Figure \(\PageIndex{6}\)). When mice were injected with the live S strain, the mice died. When he injected the mice with the live R strain or the heat-killed S strain, the mice survived. But when he injected the mice with a mixture of live R strain and heat-killed S strain, the mice died. Upon isolating the live bacteria from the dead mouse, he only recovered the S strain of bacteria. When he then injected this isolated S strain into fresh mice, the mice died. Griffith concluded that something had passed from the heat-killed S strain into the live R strain and “transformed” it into the pathogenic S strain; he called this the “transforming principle.” These experiments are now famously known as Griffith’s transformation experiments.
Query \(\PageIndex{1}\)
Query \(\PageIndex{1}\)
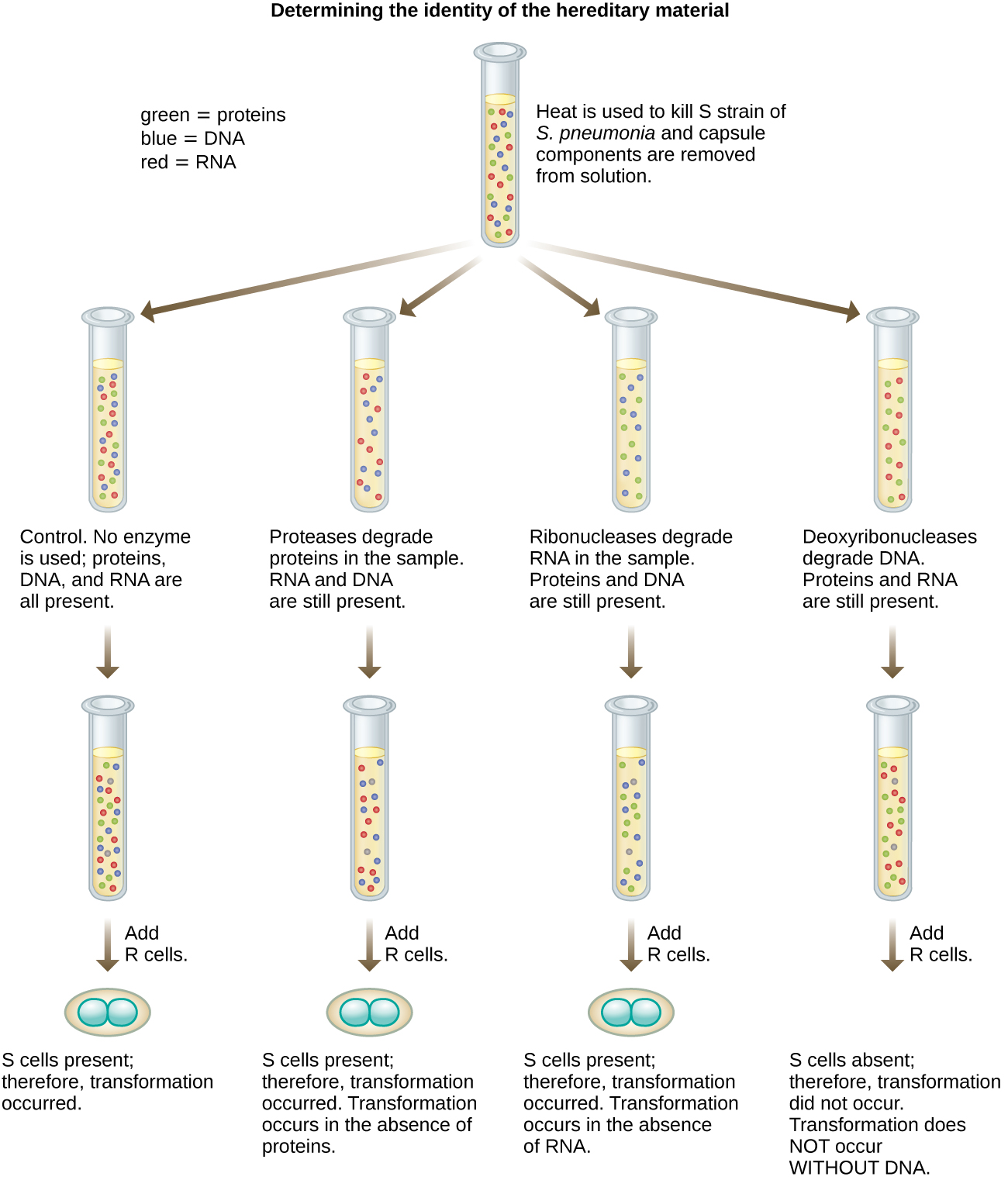
In 1944, Oswald Avery, Colin MacLeod, and Maclyn McCarty were interested in exploring Griffith’s transforming principle further. They isolated the S strain from infected dead mice, heat-killed it, and inactivated various components of the S extract, conducting a systematic elimination study (Figure \(\PageIndex{7}\)). They used enzymes that specifically degraded proteins, RNA, and DNA and mixed the S extract with each of these individual enzymes. Then, they tested each extract/enzyme combination’s resulting ability to transform the R strain, as observed by the diffuse growth of the S strain in culture media and confirmed visually by growth on plates. They found that when DNA was degraded, the resulting mixture was no longer able to transform the R strain bacteria, whereas no other enzymatic treatment was able to prevent transformation. This led them to conclude that DNA was the transforming principle. Despite their results, many scientists did not accept their conclusion, instead believing that there were protein contaminants within their extracts.
Hershey and Chase’s Proof of DNA as Genetic Material
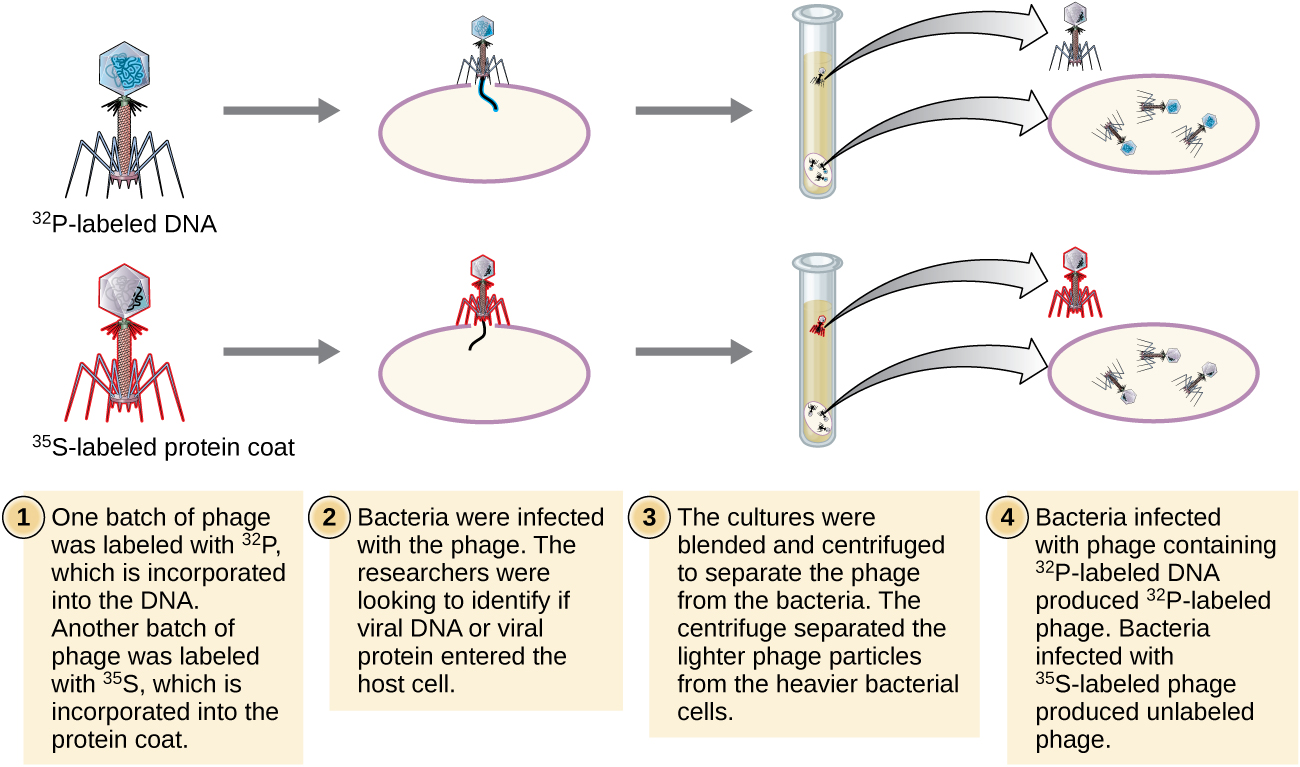
Alfred Hershey and Martha Chase performed their own experiments in 1952 and were able to provide confirmatory evidence that DNA, not protein, was the genetic material (Figure \(\PageIndex{8}\)).4 Hershey and Chase were studying a bacteriophage, a virus that infects bacteria. Viruses typically have a simple structure: a protein coat, called the capsid, and a nucleic acid core that contains the genetic material, either DNA or RNA (see Viruses). The particular bacteriophage they were studying was the T2 bacteriophage, which infects E. coli cells. As we now know today, T2 attaches to the surface of the bacterial cell and then it injects its nucleic acids inside the cell. The phage DNA makes multiple copies of itself using the host machinery, and eventually the host cell bursts, releasing a large number of bacteriophages.
Hershey and Chase labeled the protein coat in one batch of phage using radioactive sulfur, 35S, because sulfur is found in the amino acids methionine and cysteine but not in nucleic acids. They labeled the DNA in another batch using radioactive phosphorus, 32P, because phosphorus is found in DNA and RNA but not typically in protein.
Each batch of phage was allowed to infect the cells separately. After infection, Hershey and Chase put each phage bacterial suspension in a blender, which detached the phage coats from the host cell, and spun down the resulting suspension in a centrifuge. The heavier bacterial cells settled down and formed a pellet, whereas the lighter phage particles stayed in the supernatant. In the tube with the protein labeled, the radioactivity remained only in the supernatant. In the tube with the DNA labeled, the radioactivity was detected only in the bacterial cells. Hershey and Chase concluded that it was the phage DNA that was injected into the cell that carried the information to produce more phage particles, thus proving that DNA, not proteins, was the source of the genetic material. As a result of their work, the scientific community more broadly accepted DNA as the molecule responsible for heredity.
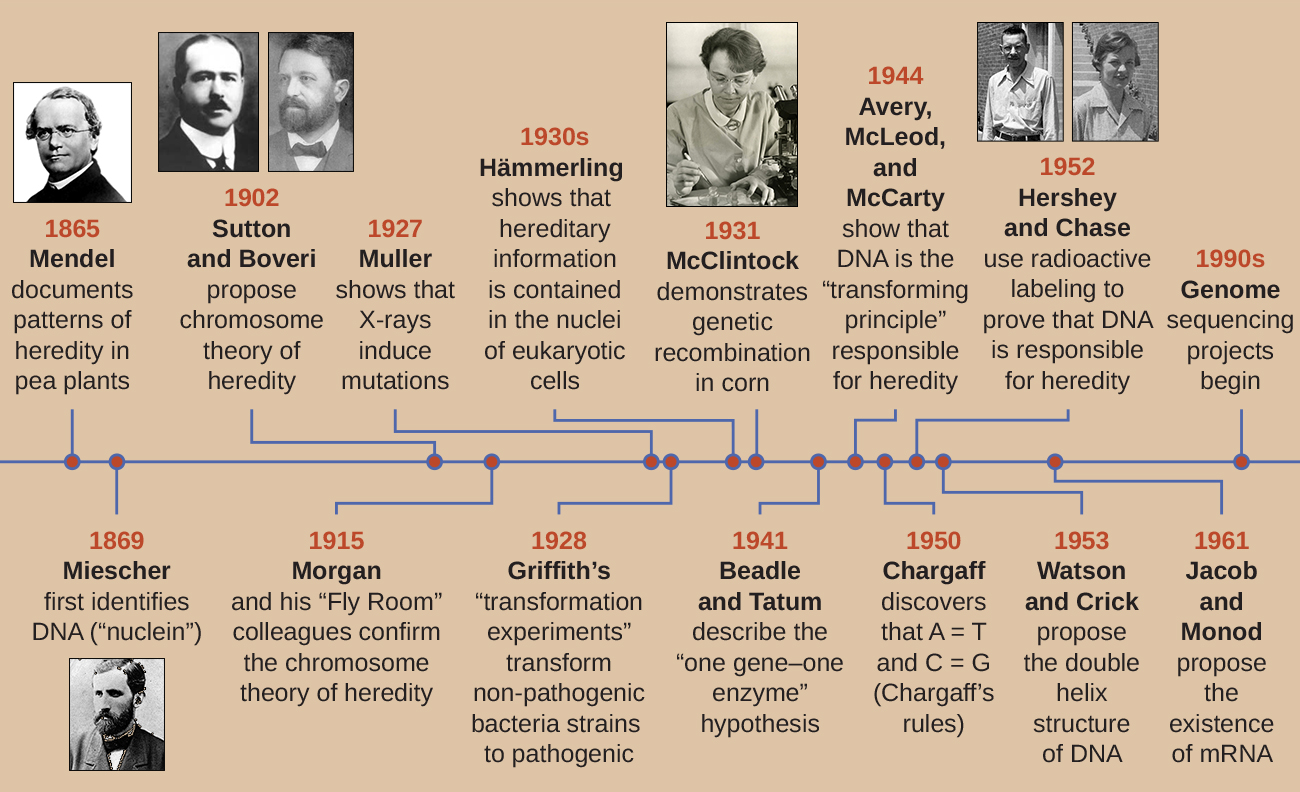
By the time Hershey and Chase published their experiment in the early 1950s, microbiologists and other scientists had been researching heredity for over 80 years. Building on one another’s research during that time culminated in the general agreement that DNA was the genetic material responsible for heredity (Figure \(\PageIndex{9}\)). This knowledge set the stage for the age of molecular biology to come and the significant advancements in biotechnology and systems biology that we are experiencing today.
Query \(\PageIndex{1}\)
To learn more about the experiments involved in the history of genetics and the discovery of DNA as the genetic material of cells, visit this website from the DNA Learning Center.
Key Concepts and Summary
- In 1928, Frederick Griffith showed that dead encapsulated bacteria could pass genetic information to live nonencapsulated bacteria and transform them into harmful strains. In 1944, Oswald Avery, Colin McLeod, and Maclyn McCarty identified the compound as DNA.
- The nature of DNA as the molecule that stores genetic information was unequivocally demonstrated in the experiment of Alfred Hershey and Martha Chase published in 1952. Labeled DNA from bacterial viruses entered and infected bacterial cells, giving rise to more viral particles. The labeled protein coats did not participate in the transmission of genetic information.
Footnotes
- 1 J.G. Mendel. “Versuche über Pflanzenhybriden.” Verhandlungen des naturforschenden Vereines in Brünn, Bd. Abhandlungen 4 (1865):3–7. (For English translation, see http://www.mendelweb.org/Mendel.plain.html)
- 2 G.W. Beadle, E.L. Tatum. “Genetic Control of Biochemical Reactions in Neurospora.” Proceedings of the National Academy of Sciences 27 no. 11 (1941):499–506.
- 3 F. Griffith. “The Significance of Pneumococcal Types.” Journal of Hygiene 27 no. 2 (1928):8–159.
- 4 A.D. Hershey, M. Chase. “Independent Functions of Viral Protein and Nucleic Acid in Growth of Bacteriophage.” Journal of General Physiology 36 no. 1 (1952):39–56.