
23.2: Cladistics
- Page ID
- 74232

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Scientists collect information that allows them to make evolutionary connections between organisms. Similar to detective work, scientists must use evidence to uncover the facts. In the case of phylogeny, evolutionary investigations focus on two types of evidence: morphologic (form and function) and genetic.
Two Measures of Similarity
Organisms that share similar physical features and genetic sequences tend to be more closely related than those that do not. Features that overlap both morphologically and genetically are referred to as homologous structures; the similarities stem from common evolutionary paths. For example, as shown in Figure \(\PageIndex{1}\), the bones in the wings of bats and birds, the arms of humans, and the foreleg of a horse are homologous structures. Notice the structure is not simply a single bone, but rather a grouping of several bones arranged in a similar way in each organism even though the elements of the structure may have changed shape and size.

Misleading Appearances
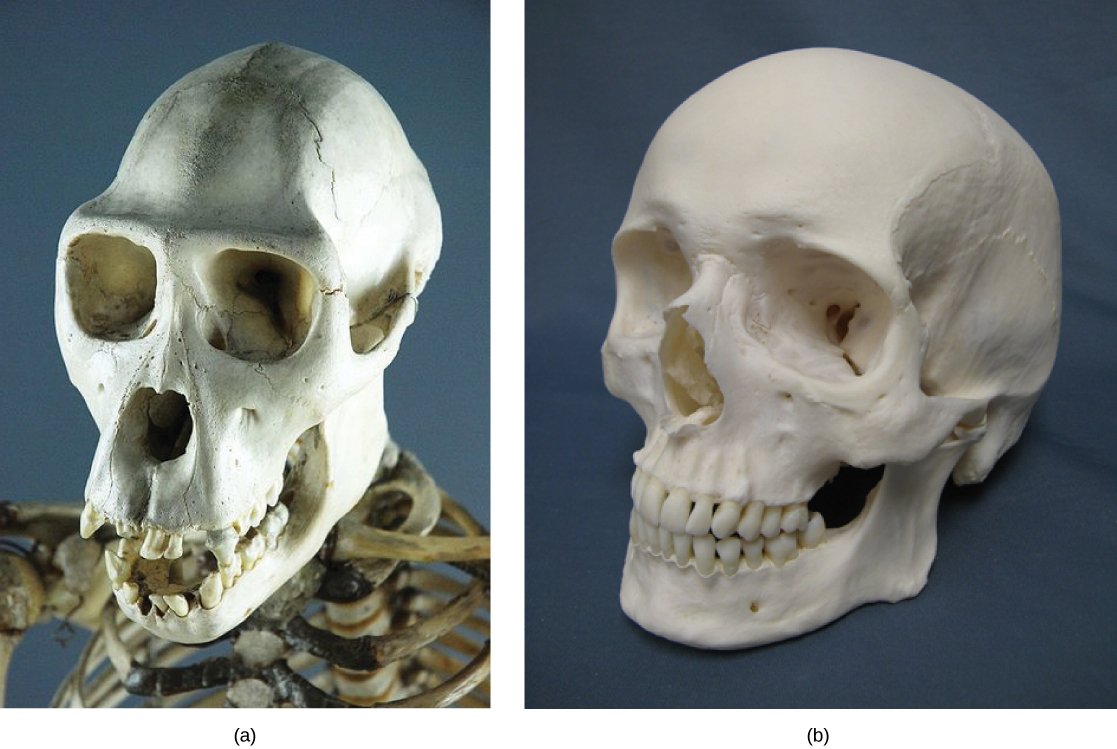
Some organisms may be very closely related, even though a minor genetic change caused a major morphological difference to make them look quite different. For example, chimpanzees and humans, the skulls of which are shown in Figure \(\PageIndex{2}\) are very similar genetically, sharing 99 percent1 of their genes. However, chimpanzees and humans show considerable anatomical differences, including the degree to which the jaw protrudes in the adult and the relative lengths of our arms and legs.
However, unrelated organisms may be distantly related yet appear very much alike, usually because common adaptations to similar environmental conditions evolved in both. An example is the streamlined body shapes, the shapes of fins and appendages, and the shape of the tails in fishes and whales, which are mammals. These structures bear superficial similarity because they are adaptations to moving and maneuvering in the same environment—water. When a characteristic that is similar occurs by adaptive convergence (convergent evolution), and not because of a close evolutionary relationship, it is called an analogous structure. In another example, insects use wings to fly like bats and birds. We call them both wings because they perform the same function and have a superficially similar form, but the embryonic origin of the two wings is completely different. The difference in the development, or embryogenesis, of the wings in each case is a signal that insects and bats or birds do not share a common ancestor that had a wing. The wing structures, shown in Figure \(\PageIndex{3}\) evolved independently in the two lineages.
Similar traits can be either homologous or analogous. Homologous traits share an evolutionary path that led to the development of that trait, and analogous traits do not. Scientists must determine which type of similarity a feature exhibits to decipher the phylogeny of the organisms being studied.

Molecular Comparisons
With the advancement of DNA technology, the area of molecular systematics, which describes the use of information on the molecular level including DNA sequencing, has blossomed. New analysis of molecular characters not only confirms many earlier classifications, but also uncovers previously made errors. Molecular characters can include differences in the amino-acid sequence of a protein, differences in the individual nucleotide sequence of a gene, or differences in the arrangements of genes. Phylogenies based on molecular characters assume that the more similar the sequences are in two organisms, the more closely related they are. Different genes change evolutionarily at different rates and this affects the level at which they are useful at identifying relationships. Rapidly evolving sequences are useful for determining the relationships among closely related species. More slowly evolving sequences are useful for determining the relationships between distantly related species. To determine the relationships between very different species such as Eukarya and Archaea, the genes used must be very ancient, slowly evolving genes that are present in both groups, such as the genes for ribosomal RNA. Comparing phylogenetic trees using different sequences and finding them similar helps to build confidence in the inferred relationships.
Sometimes two segments of DNA in distantly related organisms randomly share a high percentage of bases in the same locations, causing these organisms to appear closely related when they are not. For example, the fruit fly shares 60 percent of its DNA with humans.2 In this situation, computer-based statistical algorithms have been developed to help identify the actual relationships, and ultimately, the coupled use of both morphologic and molecular information is more effective in determining phylogeny.
EVOLUTION IN ACTION: Why Does Phylogeny Matter?
In addition to enhancing our understanding of the evolutionary history of species, our own included, phylogenetic analysis has numerous practical applications. Two of those applications include understanding the evolution and transmission of disease and making decisions about conservation efforts. A 2010 study3 of MRSA (methicillin-resistant Staphylococcus aureus), an antibiotic resistant pathogenic bacterium, traced the origin and spread of the strain throughout the past 40 years. The study uncovered the timing and patterns in which the resistant strain moved from its point of origin in Europe to centers of infection and evolution in South America, Asia, North America, and Australasia. The study suggested that introductions of the bacteria to new populations occurred very few times, perhaps only once, and then spread from that limited number of individuals. This is in contrast to the possibility that many individuals had carried the bacteria from one place to another. This result suggests that public health officials should concentrate on quickly identifying the contacts of individuals infected with a new strain of bacteria to control its spread.
A second area of usefulness for phylogenetic analysis is in conservation. Biologists have argued that it is important to protect species throughout a phylogenetic tree rather than just those from one branch of the tree. Doing this will preserve more of the variation produced by evolution. For example, conservation efforts should focus on a single species without sister species rather than another species that has a cluster of close sister species that recently evolved. If the single evolutionarily distinct species goes extinct a disproportionate amount of variation from the tree will be lost compared to one species in the cluster of closely related species. A study published in 20074 made recommendations for conservation of mammal species worldwide based on how evolutionarily distinct and at risk of extinction they are. The study found that their recommendations differed from priorities based on simply the level of extinction threat to the species. The study recommended protecting some threatened and valued large mammals such as the orangutans, the giant and lesser pandas, and the African and Asian elephants. But they also found that some much lesser known species should be protected based on how evolutionary distinct they are. These include a number of rodents, bats, shrews and hedgehogs. In addition there are some critically endangered species that did not rate as very important in evolutionary distinctiveness including species of deer mice and gerbils. While many criteria affect conservation decisions, preserving phylogenetic diversity provides an objective way to protect the full range of diversity generated by evolution.
Building Phylogenetic Trees
How do scientists construct phylogenetic trees? Presently, the most accepted method for constructing phylogenetic trees is a method called cladistics. This method sorts organisms into clades, groups of organisms that are most closely related to each other and the ancestor from which they descended. For example, in Figure \(\PageIndex{4}\), all of the organisms in the shaded region evolved from a single ancestor that had amniotic eggs. Consequently, all of these organisms also have amniotic eggs and make a single clade, also called a monophyletic group. Clades must include the ancestral species and all of the descendants from a branch point.
ART CONNECTION
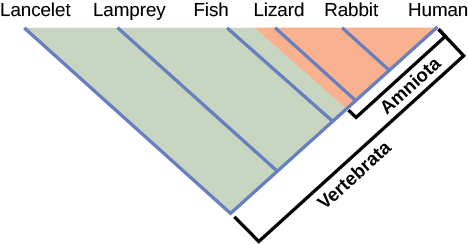
Which animals in this figure belong to a clade that includes animals with hair? Which evolved first: hair or the amniotic egg?
Clades can vary in size depending on which branch point is being referenced. The important factor is that all of the organisms in the clade or monophyletic group stem from a single point on the tree. This can be remembered because monophyletic breaks down into “mono,” meaning one, and “phyletic,” meaning evolutionary relationship.
Shared Characteristics
Cladistics rests on three assumptions. The first is that living things are related by descent from a common ancestor, which is a general assumption of evolution. The second is that speciation occurs by splits of one species into two, never more than two at a time, and essentially at one point in time. This is somewhat controversial, but is acceptable to most biologists as a simplification. The third assumption is that traits change enough over time to be considered to be in a different state .It is also assumed that one can identify the actual direction of change for a state. In other words, we assume that an amniotic egg is a later character state than non-amniotic eggs. This is called the polarity of the character change. We know this by reference to a group outside the clade: for example, insects have non-amniotic eggs; therefore, this is the older or ancestral character state. Cladistics compares ingroups and outgroups. An ingroup (lizard, rabbit and human in our example) is the group of taxa being analyzed. An outgroup (lancelet, lamprey and fish in our example) is a species or group of species that diverged before the lineage containing the group(s) of interest. By comparing ingroup members to each other and to the outgroup members, we can determine which characteristics are evolutionary modifications determining the branch points of the ingroup’s phylogeny.
If a characteristic is found in all of the members of a group, it is a shared ancestral characterbecause there has been no change in the trait during the descent of each of the members of the clade. Although these traits appear interesting because they unify the clade, in cladistics they are considered not helpful when we are trying to determine the relationships of the members of the clade because every member is the same. In contrast, consider the amniotic egg characteristic of Figure \(\PageIndex{4}\). Only some of the organisms have this trait, and to those that do, it is called a shared derived character because this trait changed at some point during descent. This character does tell us about the relationships among the members of the clade; it tells us that lizards, rabbits, and humans group more closely together than any of these organisms do with fish, lampreys, and lancelets.
A sometimes confusing aspect of “ancestral” and “derived” characters is that these terms are relative. The same trait could be either ancestral or derived depending on the diagram being used and the organisms being compared. Scientists find these terms useful when distinguishing between clades during the building of phylogenetic trees, but it is important to remember that their meaning depends on context.
Choosing the Right Relationships
Constructing a phylogenetic tree, or cladogram, from the character data is a monumental task that is usually left up to a computer. The computer draws a tree such that all of the clades share the same list of derived characters. But there are other decisions to be made, for example, what if a species presence in a clade is supported by all of the shared derived characters for that clade except one? One conclusion is that the trait evolved in the ancestor, but then changed back in that one species. Also a character state that appears in two clades must be assumed to have evolved independently in those clades. These inconsistencies are common in trees drawn from character data and complicate the decision-making process about which tree most closely represents the real relationships among the taxa.
To aid in the tremendous task of choosing the best tree, scientists often use a concept called maximum parsimony, which means that events occurred in the simplest, most obvious way. This means that the “best” tree is the one with the fewest number of character reversals, the fewest number of independent character changes, and the fewest number of character changes throughout the tree. Computer programs search through all of the possible trees to find the small number of trees with the simplest evolutionary pathways. Starting with all of the homologous traits in a group of organisms, scientists can determine the order of evolutionary events of which those traits occurred that is the most obvious and simple.
CONCEPT IN ACTION

Practice Parsimony: Go to this website to learn how maximum parsimony is used to create phylogenetic trees (be sure to continue to the second page).
These tools and concepts are only a few of the strategies scientists use to tackle the task of revealing the evolutionary history of life on Earth. Recently, newer technologies have uncovered surprising discoveries with unexpected relationships, such as the fact that people seem to be more closely related to fungi than fungi are to plants. Sound unbelievable? As the information about DNA sequences grows, scientists will become closer to mapping the evolutionary history of all life on Earth.
Section Summary
To build phylogenetic trees, scientists must collect character information that allows them to make evolutionary connections between organisms. Using morphologic and molecular data, scientists work to identify homologous characteristics and genes. Similarities between organisms can stem either from shared evolutionary history (homologies) or from separate evolutionary paths (analogies). After homologous information is identified, scientists use cladistics to organize these events as a means to determine an evolutionary timeline. Scientists apply the concept of maximum parsimony, which states that the likeliest order of events is probably the simplest shortest path. For evolutionary events, this would be the path with the least number of major divergences that correlate with the evidence.
Art Connections
Figure \(\PageIndex{3}\): Which animals in this figure belong to a clade that includes animals with hair? Which evolved first: hair or the amniotic egg?
- Answer
-
Rabbits and humans belong in the clade that includes animals with hair. The amniotic egg evolved before hair, because the Amniota clade branches off earlier than the clade that encompasses animals with hair.
Footnotes
- 1 Gibbons, A. (2012, June 13). Science Now. Retrieved from news.sciencemag.org/scienceno...sequenced.html
- 2 Background on comparative genomic analysis. (2002, December). Retrieved from http://www.genome.gov/10005835
- 3 Harris, S.R. et al. 2010. Evolution of MRSA during hospital transmission and intercontinental spread. Science 327:469–474.
- 4 Isaac NJ, Turvey ST, Collen B, Waterman C, Baillie JE (2007) Mammals on the EDGE: Conservation Priorities Based on Threat and Phylogeny. PLoS ONE 2(3): e296. doi:10.1371/journal.pone.0000296
Glossary
- analogous structure
- a character found in two taxa that looks similar because of convergent evolution, not because of descent from a common ancestor
- clade
- a group of taxa with the same set of shared derived characters, including an ancestral species and all its descendants
- cladistics
- a method used to organize homologous traits to describe phylogenies using common descendent as the primary criterion used to classify organisms
- maximum parsimony
- applying the simplest, most obvious way with the least number of steps
- molecular systematics
- the methods of using molecular evidence to identify phylogenetic relationships
- monophyletic group
- (also, clade) organisms that share a single ancestor
- shared ancestral character
- a character on a phylogenetic branch that is shared by a particular clade
- shared derived character
- a character on a phylogenetic tree that is shared only by a certain clade of organisms
Contributors and Attributions
Samantha Fowler (Clayton State University), Rebecca Roush (Sandhills Community College), James Wise (Hampton University). Original content by OpenStax (CC BY 4.0; Access for free at https://cnx.org/contents/b3c1e1d2-83...4-e119a8aafbdd).