
9.1: Transposable Elements (Transposons)
- Page ID
- 374

Transposable elements (both active and inactive) occupy approximately half the human genome and a substantially greater fraction of some plant genomes! These movable elements are ubiquitous in the biosphere, and are highly successful in propagating themselves. We now realize that some transposable elements are also viruses, for instance, some retroviruses can integrate into a host genome to form endogenous retroviruses. Indeed, some viruses may be derived from natural transposable elements and vice versa. Since viruses move between individuals, at least some transposable elements can move between genomes (between individuals) as well as within an individual’s genome. Given their prevalence in genomes, the function (if any) of transposable elements has been much discussed but is little understood. It is not even clear whether transposable elements should be considered an integral part of a species’ genome, or if they are successful parasites. They do have important effects on genes and their phenotypes, and they are the subject of intense investigation.
Transposition is related to replication, recombination and repair. The process of moving from one place to another involves a type of recombination, insertions of transposable elements can cause mutations, and some transpositions are replicative, generating a new copy while leaving the old copy intact. However, this ability to move is a unique property of transposable elements, and warrants treatment by itself.
Properties and effects of transposable elements
The defining property of transposable elements is their mobility; i.e. they are genetic elements that can move from one position to another in the genome. Beyond the common property of mobility, transposable elements show considerable diversity. Some move by DNA intermediates, and others move by RNA intermediates. Much of the mechanism of transposition is distinctive for these two classes, but all transposable elements effectively insert at staggered breaks in chromosomes. Some transposable elements move in a replicative manner, whereas others are nonreplicative, i.e. they move without making a copy of themselves.
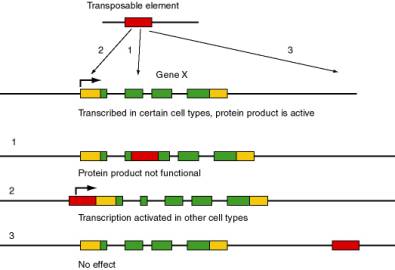
Transposable elements are major forces in the evolution and rearrangement of genomes (Figure 9.1). Some transposition events inactivate genes, since the coding potential or expression of a gene is disrupted by insertion of the transposable element. A classic example is the r allele (rugosus) of the gene encoding a starch branching enzyme in peas is nonfunctional due to the insertion of a transposable element. This allele causes the wrinkled pea phenotype in homozygotes originally studied by Mendel. In other cases, transposition can activate nearby genes by bringing an enhancer of transcription (within the transposable element) close enough to a gene to stimulate its expression. If the target gene is not usually expressed in a certain cell type, this activation can lead to pathology, such as activation of a proto-oncogene causing a cell to become cancerous. In other cases, no obvious phenotype results from the transposition. A particular type of transposable element can activate, inactivate or have no effect on nearby genes, depending on exactly where it inserts, it’s orientation and other factors.
Transposable elements can cause deletions or inversions of DNA. When transposition generates two copies of the same sequence in the same orientation, recombination can delete the DNA between them. If the two copies are in the opposite orientations, recombination will invert the DNA between them.
As part of the mechanism of transposition, additional DNA sequences can be mobilized. DNA located between two copies of a transposable element can be moved together with them when they move. In this manner, transposition can move DNA sequences that are not normally part of a transposable element to new locations. Indeed, "host" sequences can be acquired by viruses and propagated by infection of other individuals. This may be a natural means for evolving new strains of viruses. One of the most striking examples is the acquisition and modification of a proto-oncogene, such as cellular c-src, by a retrovirus to generate a modified, transforming form of the gene, called v-src. These and related observations provided insights into the progression of events that turn a normal cell into a cancerous one. They also point to the continual acquisition (and possibly deletion) of information from host genomes as a natural part of the evolution of viruses.