
Linear Templates
- Page ID
- 331

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)The problem of linear templates
The requirement of DNA polymerases to have a primer causes a problem at the ends of linear templates. As illustrated in Figure 6.13, leading strand synthesis can proceed to the end of its template strand, but lagging strand synthesis cannot. As lagging strand synthesis nears the end of its template, at some point no binding site will be available for primase, and part of the 3’ end of the template for lagging strand synthesis will not be copied. Hence a 3’ overhang is left after the replication fork has finished, and part of the chromosome is not copied into new DNA. If nothing else were done, the chromosome would become progressively shorter after each round of replication.
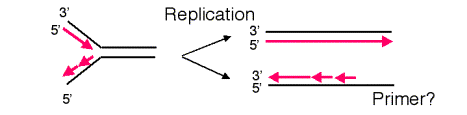
At least three different types of solution to this problem have been discovered in various organisms. One, utilized by bacteriophage such as l and T4, is to convert the linear template to a circle. For instance, the linear chromosome of bacteriophage l has cohesive ends (complementary single strands at each end, generated by a phage endonuclease) that can anneal upon infection, thereby forming a circular template for replication. Other viruses, such as adenovirus, attach a protein to the end of unreplicated DNA to serve as a primer. Such an attached protein obviates the requirement for using the unreplicated DNA as a template, and the entire viral chromosome can be replicated.
A third solution is to make the ends a series of simple repeats that are synthesized in a process distinct from DNA replication. Indeed, the ends of the linear chromosomes of most (perhaps all) eukaryotes, called telomeres, are composed of many copies of a simple repetitive sequence. This sequence is distinctive for different organisms, but in all cases one strand is rich in G and the other is rich in C. The repeating unit for human telomeres is 5' AGGGTT 3' running from the centromeric end of the repeats to the telomeric end), and the repeating units for the ciliate Tetrahymena is 5' GGGGTT 3’.
New copies of the telomeric repeats can be synthesized each time the chromosome replicates (Figure 6.14). This re-synthesis of the telomeric repeats counteracts the progressive shortening of the linear chromosomes that would occur if only the replication forks were used to synthesize new chromosomes.
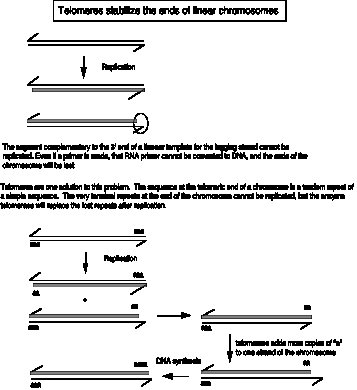
In this figure, the string of "a" at the ends of the chromosome is the tandem repeat of simple sequence, in duplex form. For instance, for a human chromosome, "a" would be
CEN ... 5' AGGGTT 3' ... TEL
3' TCCCAA 5'
or for a Tetrahymena chromosome, "a" would be
CEN ... 5' GGGGTT 3' ... TEL
3' CCCCAA 5'
In each case, the "a" or monomer is repeated thousands of times in tandem.
Addition of new telomeric repeats is catalyzed by the enzyme telomerase. As illustrated in Figure 6.15, this enzyme catalyzes many successive rounds of synthesis, adding many copies of the simple repeat to the ends of the chromosomes. The enzyme is a ribonucleoprotein, i.e. it has both a polypeptide and an RNA component. The RNA serves as a template to direct addition of nucleotides to the 3' end of the G+T rich strand, and the polypeptide acts as a reverse transcriptase to make a DNA copy of a hexanuclotide segment of the RNA. For instance, the telomerase from Tetrahymena will copy the 3’CCCCAA in the RNA template into 5’GGGGTT telomeric repeat. Then the enzyme shifts over and synthesizes another hexanucleotide. The fact that the RNA serves as the template was demonstrated by exchanging the RNA component of isolated telomerase with the telomerase RNA from a second species. This exchange led to the addition of telomeres with sequences characteristic of that of the second species, showing that the telomerase RNA is the determinant of the sequence of the telomere. The protein component provides the reverse transcriptase activity.
Once many copies of the G+T-rich strand of telomeres have been synthesized by telomerase, the long single strand forms a specialized structure toward the 3’ end. Some evidence indicates that a “G-quartet” is formed, in which four guanine nucleotides form a hydrogen-bonded complex. Examination of the ends of replicating chromosomes in the electron microscope show a circular structure. Although details of the structure at the end of this strand are not fully established, it is likely that a primer to support synthesis of the C+A-rich strand is made effectively by turning the G+T-rich strand around. Conventional synthesis by DNA polymerases can then copy the G+T-rich strand to make the complementary strand. Some processing, e.g. nucleases acting at the end, can convert the specialized structure or hairpin into a linear duplex.

How processive is telomerase?
Not all replicating cells have telomerase activity. This activity is higher in some transformed cells than in nontransformed cells. Also, older cells tend to have shorter telomeres. Thus telomeres are being actively investigated as possibly playing roles in both aging and in tumorigenic transformation.
Telomeres are important for stabilizing chromosomes. Some chromosomal deletions remove the ends of the chromosome, including the telomere, and these shortened chromosomes are less stable than their wild-type counterparts. Directed mutations have been made in mice to eliminate telomerase activity. These mice are viable for several generations, but they show many broken and abnormal chromosomes, demonstrating the importance of this activity.