
10.1: Plant Diversity Imbalance
- Page ID
- 21635

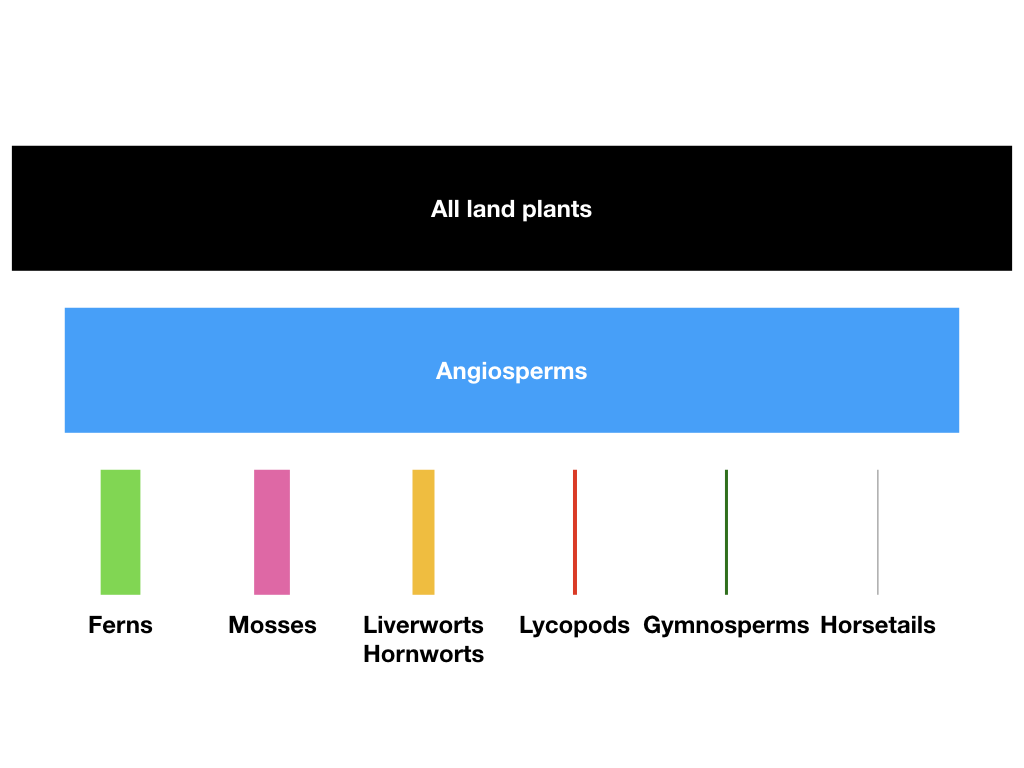
The diversity of flowering plants (the angiosperms) dwarfs the number of species of their closest evolutionary relatives (Figure 10.1). There are more than 260,000 species of angiosperms (that we know; more are added every day). The clade originated more than 140 million years ago (Bell et al. 2005), and all of these species have formed since then. One can contrast the diversity of angiosperms with the diversity of other groups that originated at around the same time. For example, gymnosperms, which are as old as angiosperms, include only around 1000 species, and may even represent more than one clade. The diversity of angiosperms also dwarfs the diversity of familiar vertebrate groups of similar age (e.g. squamates - snakes and lizards - which diverged from their sister taxon, the tuatara, some 250 mya or more Hedges et al. 2006, include fewer than 8000 species).
The evolutionary rise of angiosperm diversity puzzled Darwin over his career, and the issues surrounding angiosperm diversification are often referred to as “Darwin’s abominable mystery” in the scientific literature (e.g. Davies et al. 2004). The main mystery is the tremendous variation in numbers of species across plant clades (see Figure 10.1). This variation even applies within angiosperms, where some clades are much more diverse than others.
At a global scale, the number of species in a clade can change only via two processes: speciation and extinction. This means that we must look to speciation and extinction rates – and how they vary through time and across clades – to explain phenomena like the extraordinary diversity of Angiosperms. It is to this topic that we turn in the next few chapters. Since Darwin’s time, we have learned a lot about the evolutionary processes that led to the diversity of angiosperms that we see today. These data provide an incredible window into the causes and effects of speciation and extinction over macroevolutionary time scales.
Comparative methods can be applied to understand patterns of species richness by estimating speciation and extinction rates, both across clades and through time. In this chapter, I will introduce birth-death models, by far the most common model for understanding diversification in a comparative framework. I will discuss the mathematics of birth-death models and how these models relate to the shapes of phylogenetic trees. I will describe how to simulate phylogenetic trees under a birth-death model. Finally, I will discuss tree balance and lineage-through-time plots, two common ways to measure the shapes of phylogenetic trees.