
18.3: The Molecular Structure and Sub-Cellular Organization of Cytoskeletal Components
- Page ID
- 16529

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Of the three main cytoskeletal fibers, intermediate filaments serve a mainly structural role in cells. Microtubules and microfilaments have dual functions, dynamically maintaining cell shape and enabling cell motility. For example, when attached to the plasma membrane, microfilaments maintain cell shape. However, by interacting with motor proteins (e.g., myosin), they can pull or push against a muscle cell membrane, changing the shape of the cell. Likewise, motor proteins such as dynein and kinesin can move ‘cargo’ to and fro along microtubule tracks from one point to another in the cell. We will look at how motor proteins interact with microtubules and microfilaments shortly. At this point, let’s take another look at the drawings and micrographs of the three main cytoskeletal filaments of eukaryotic cells (below) that we saw earlier in the text.
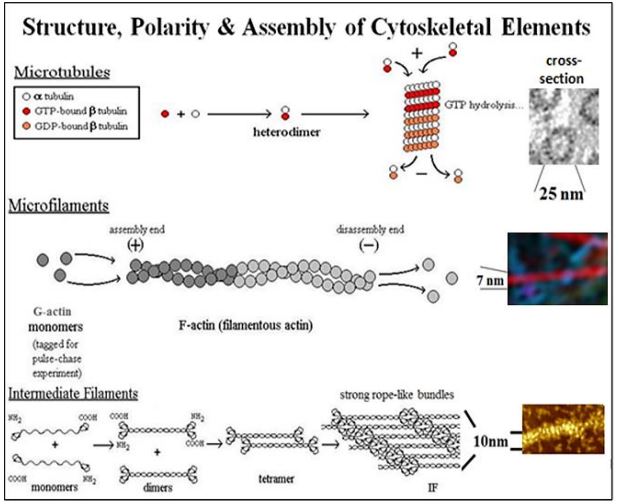
321 Introduction to the Cytoskeleton
The location and general functions of microtubules, microfilaments and intermediate filaments were demonstrated by immunofluorescence microscopy. After exposing cells to fluorophore-tagged antibodies against either microtubule, microfilament (actin) or intermediate filament proteins, fluorescence micrographs of the stained cells revealed the different locations of the fibers in cells. The typical localization of the different cytoskeletal fibers is shown below.

These localizations are consistent with known functions of the major cytoskeletal component filaments in cell structure and motility. Despite the small size of prokaryotic cells, they too were recently found have previously unsuspected cytoplasmic structures that could serve as a cytoskeleton (ncbi-A Prokaryotic Cytoskeleton?). So perhaps all (not just eukaryotic) cells are more than an unorganized bag of fluid sap! Next, we consider specific roles of microtubules, microfilaments, intermediate filaments and related proteins in the eukaryotic cytoskeleton.
322 Microtubules, Microfilaments, and Intermediate Filaments in Cells
A. Microtubules - an Overview
Microtubules assemble from dimers of \(\alpha \)-tubulin and \(\beta \)-tubulin monomers. After formation, \(\alpha \)/\(\beta \)-tubulin dimers add to a growing, or plus end (+end), fueled by GTP hydrolysis. Disassembly at the -end of microtubules powers changing the shape of cells or the separation and movement of chromatids to opposite poles of cells during cell division (i.e., mitosis or meiosis). Isolated single microtubules were shown to grow by addition to one end and to disassemble at the opposite end, thus distinguishing the +ends and -ends. A summary of this experiment demonstrating microtubule polarity is in the link below.
323 Demonstration of the Polarity and Dynamics of Microtubules
Microtubules in most cells can seem disordered. In interphase, they tend to radiate from centrioles in non-dividing animal cells, without forming discrete structures. However, in the run-up to cell division, microtubules reorganize to form spindle fibers. This reorganization is nucleated from centrioles in animal cells and from a more amorphous microtubule organizing center (MTOC) in plant cells. A typical centriole (or basal body) in has a ‘9 triplet’ microtubule array as seen in the electron micrograph cross section (below).
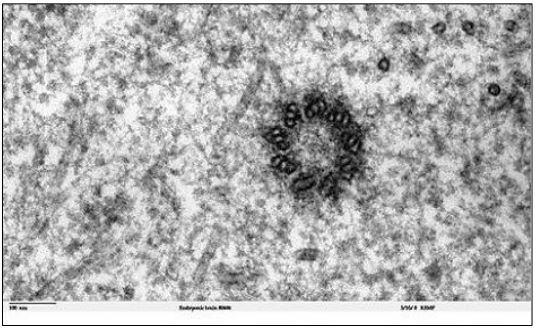
1. The Two Kinds of Microtubules in Spindle Fibers
a) Kinetochore Microtubules
Duplicated chromosomes condense in prophase of mitosis and meiosis, forming visible paired chromatids attached at their centromeres. Specific proteins associate with centromeres to make a kinetochore during condensation. As the spindle apparatus forms, some spindle fibers attach to the kinetochore; these are the kinetochore microtubules. By metaphase, bundles of kinetochore microtubules stretch from the kinetochores at the cell center to the polar centrioles or MTOCs of the dividing cell, as drawn below.

We now know that the +ends of kinetochore microtubules are in fact at the kinetochores, where these fibers are assembled! At anaphase, forces generated when microtubules shorten at their –ends (disassembly ends) separate the chromatids. Microtubule disassembly at centrioles/MTOCs provides the force that draws daughter chromosomes to the opposite poles of the cell as cell division continues.
b) Polar Microtubules
The spindle fiber polar microtubules extend from centrioles/MTOCs at opposite poles of the cell. They do not bind to kinetochores of chromatids, but instead, overlap at the center of the dividing cells. As kinetochore microtubules pull at chromatids in anaphase, polar microtubules slide past one another in opposite directions, pushing apart the poles of the cell. In this case, dynein (a motor protein attached to microtubules) catalyzes ATP hydrolysis to power microtubule sliding. Dynein motors on the microtubules from one pole of the cell in effect, ‘walk’ along overlapping microtubules extending from the opposite pole. The role of microtubule disassembly at the centrioles (i.e., at the minus end) was demonstrated in a clever experiment in which a tiny laser beam was aimed into a cell at spindle fibers attached to the kinetochore of a pair of chromatids (see this animated at the link below).
2. Microtubules in Cilia and Flagella
The microtubules of cilia or flagella emerge from a basal body, shown at the left in the electron micrograph below
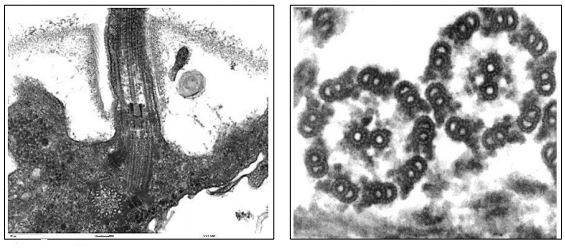
Basal bodies are structurally similar to centrioles, organized as a ring of nine microtubule triplets. Cilia and flagella formation begin at basal bodies but show a typical 9+2 arrangement (9 outer doublet plus 2 central microtubules) in cross section (shown in the micrograph, above right). After detergent treatment to remove the membranes of isolated cilia or flagella, the remaining axonemes preserve the 9+2 microtubule arrangement. The structural relationship between the axonemes of a cilium or flagellum and an individual microtubule are shown in the cross sections below.

It is possible to see the tubulin subunits that make up a microtubule polymer in cross section. Each tubule is made up of a ring of 13 tubulin subunits. The microtubules in the ‘doublets’ share tubulins, but are also composed of 13 tubulins. When fully formed, the 25 nm diameter microtubules appear to be a hollow cylinder. When microtubules are isolated, they typically come along with dynein motor proteins and other Microtubule-Associated Proteins (MAPs), some of which hold microtubules together in an axoneme.
3. Microtubule Motor Proteins Move Cargo from Place to Place in Cells
Motor proteins such as dynein and kinesin, are ATPases; they use the free energy of ATP hydrolysis to power intracellular motility. Let’s take a closer look at how these two major motor proteins carry cargo from place to place inside of cells. Organelles are a typical cargo. Examples include vesicles formed at the trans Golgi face containing secretory proteins, pigments or neurotransmitters. Secretory vesicles move along microtubule tracks to the plasma membrane for exocytosis. Vesicles containing neurotransmitters move from the cell body of neurons along microtubule tracks in the axons, reaching the nerve ending where they become synaptic vesicles. In a chameleon, pigment vesicles in skin cells disperse or aggregate along microtubule tracks to change skin color to match the background.
Motor proteins carry cargo vesicles in opposite directions. The transport of neurotransmitters is a well-understood example. Neurotransmitter vesicles arise from the endomembrane system in neuron cell bodies. Powered by ATP, kinesin powers anterograde vesicle movement from the cell body to nerve endings. In contrast, an ATP-dependent dynein motor, as part of a dynactin complex, powers retrograde movement of empty vesicles back to the cell body. Motor protein structure and action are below.
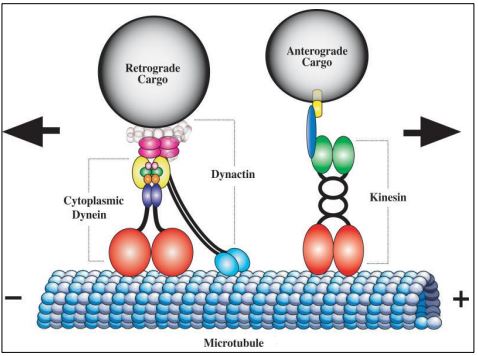
A fanciful (and not too inaccurate!) cartoon of a motor protein ‘walking along an axonal microtubule is animated at this link: Kinesin 'walking' an organelle along a microtubule.
325 Microtubule Motor Proteins
At this point, we can look at several specific kinds of cell motility involving microtubules and microfilaments.
4. The Motor Protein Dynein Enables Axonemes to Bend
Take a look at the cross-section of axonemes a few illustrations ago. In the 9+2 axoneme of cilia and flagella, dynein arms attached to the A tubules of the outer doublets walk along the B tubules of the adjacent doublet. If only the doublets on one side of an axoneme take a walk while those on the other side hold still, the microtubules will slide past one another and the axoneme (and therefore a cilium or flagellum) will bend. However, microtubule sliding is constrained by flexible nexin and radial spoke attachments. The movements of cilia and flagella are illustrated below.

The differences in flagellar motion (wave-like propeller) and ciliary motion (single plane, back and forth beat) result in part from which microtubules are sliding at a given moment and the nature of their restraint by axoneme proteins. Let’s look at some experiments that demonstrate these events. CC-BY 3.0; From: en.Wikipedia.org/wiki/File:Fl...um-beating.svg CMB3e 467.
Experiments on isolated axonemes demonstrate the sliding microtubule mechanism of ciliary and flagellar motility. In one experiment, isolated flagella and purified axonemes were both shown to ‘beat’ in the presence of added ATP (below).

Agitating sperm or ciliated cells in a high-speed blender for a few seconds will shear and detach flagella or cilia from the rest of the cell. Adding ATP to detached cilia or flagella will cause them to beat, a phenomenon easily seen in a light microscope. Axonemes isolated from detached cilia or flagella by detergent treatment (to disrupt membranes) retain their characteristic 9+2 microtubule arrangement as well as other ultrastructural features…, and will even ‘beat’ in the presence of ATP!
326 9+2 Microtubule Array in Axonemes that Beat
Additional detergent treatment removes radial spokes, nexin and other proteins from the axoneme, causing the microtubules to separate. Dissociated microtubule doublets and central ‘singlets’ can then be observed in the electron microscope.
When separated microtubules are dialyzed to remove the detergents, the doublet microtubules re-associate, forming sheets, as shown in the cartoon below.

ATP added to these ‘reconstituted’ microtubule doublets causes the microtubules to separate as the ATP is hydrolyzed. When such preparations are fixed for electron microscopy immediately after adding the ATP, they are caught in the act of sliding. See this animated in the first link below.
327 Proof of Sliding Microtubules During the Bending of Flagella and Cilia
328 Bacterial Flagella are Powered by a Proton Gradient
329 The Effects of Different Drugs on Microtubules and Cancer
C. Microfilaments - Structure and Role in Muscle Contraction
At 7 nm in diameter, microfilaments (actin filaments) are the thinnest cytoskeletal component. Globular actin (G-actin) monomers polymerize to form linear F-actin polymers. Two polymers then combine to form the twin-helical actin microfilament. As with microtubules, microfilaments have a +end to which new actin monomers are added to assemble F-actin, and a –end at which they disassemble when they are in a dynamic state, such as when a cell is changing shape. When one end of a microfilament is anchored to a cellular structure, for example to plaques in the cell membrane, motor proteins like myosin can use ATP to generate a force that deforms the plasma membrane and thus, the shape of the cell. One of the best-studied examples of myosin/actin interaction is in skeletal muscle where the sliding of highly organized thick myosin rods and the thin actin microfilaments results in muscle contraction.
1. Thick and Thin Filaments of Skeletal Muscle Contraction
Bundles of parallel muscle cells make up a skeletal muscle. Light microscopy of skeletal muscle thin sections show striated muscle cells (myocytes, below).
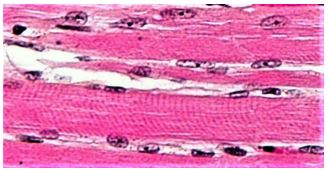
The dark purplish structures surrounding the myocyte are mitochondria, which will provide the ATP to fuel contraction. Skeletal muscle is made up of ‘aligned’, bundled myocytes. The bundled myocytes (also called myofibers) are further organized into fascicles that are finally bundled into a muscle. The blowout illustration on the next page shows this anatomical organization and fine structure of a muscle (left panel).
High-resolution electron microscopy from the 1940s revealed the fine structure of skeletal muscle (right panel of the illustration), allowing characterization of the sarcomere. The dark bands of the striations in the light micrograph of myocytes are regions of aligned, adjacent sarcomeres. A pair of Z lines demarcate a sarcomere (Z for zwischen, German for between). The I band is a relatively clear region of the sarcomere, largely made up of thin (actin) microfilaments. The A band at the center of the sarcomere consists of overlapping thin and thick (actin and myosin) filaments, while the H zone is a region where myosin does not overlap actin filaments. An M line lies at the center of the H zone. Multiple repeating sarcomeres of myocytes aligned in register in the fascicles give the appearance of striations in whole muscles.
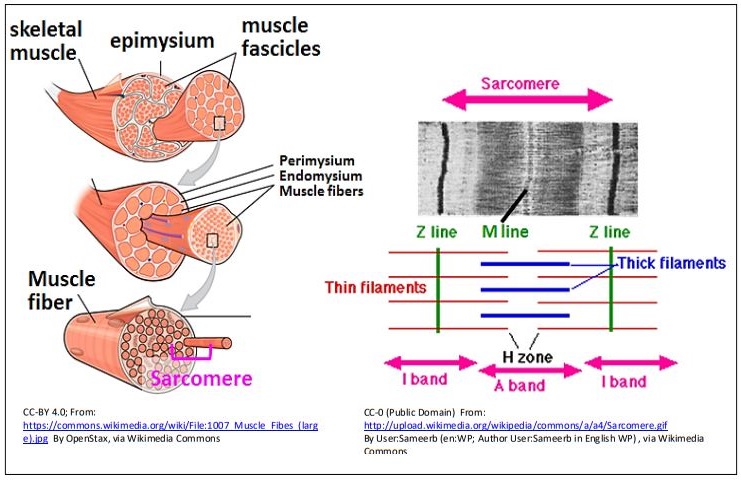
2. The Sliding Filament Model of Skeletal Muscle Contraction
Electron microscopy of relaxed and contracted muscle shown below is consistent with the sliding of thick and thin filaments during contraction. Additional key structures of the sarcomere can be seen in the drawing at the right.

Note that in the sarcomeres of a contracted muscle cell, the H zone has almost disappeared. While the width of the A band has not changed after contraction, the width of the I bands has decreased and the Z-lines are closer in the contracted sarcomere. The best explanation here was the Sliding Filament Hypothesis (model) of skeletal muscle contraction.
330 The Sliding Filament Model of Skeletal Muscle Contraction
3. The Contraction Paradox: Contraction and Relaxation Require ATP
The role of ATP in fueling the movement of sliding filaments during skeletal muscle contraction was based in part on experiments with glycerinated fibers (muscle fibers soaked in glycerin to permeabilize the plasma membrane). The soluble cytoplasmic components leak out of glycerinated fibers, but leave the sarcomere structures intact, as visualized by electron microscopy. Investigators found that, if ATP and calcium were added back to glycerinated fibers, the ATP was hydrolyzed and the fiber could still contract… and even lift a weight!
The contraction of a glycerinated muscle fiber in the presence of ATP is illustrated below.
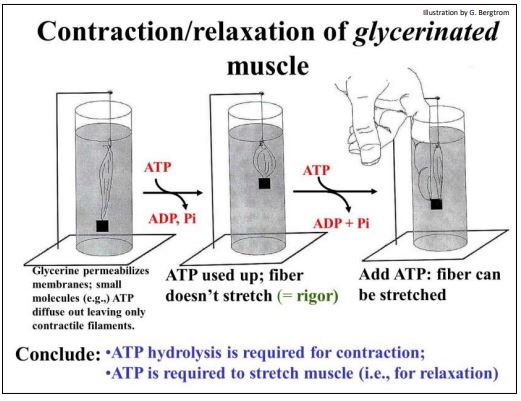
When assays showed that all of the added ATP had been hydrolyzed, the muscle remained contracted. It would not relax, even with the weight it had lifted still attached! Attempting to manually force the muscle back to its relaxed position didn’t work. But the fiber could be stretched when fresh ATP was added to the preparation! Moreover, if the experimenter let go immediately after stretching the fiber, it would again contract and lift the weight! A cycle of forced stretching and contraction could be repeated until all of the added ATP was hydrolyzed. At that point, the fiber would again no longer contract…, or if contracted, could no longer be stretched.
The contraction paradox then, was that ATP hydrolysis was required for muscle contraction as well as for relaxation (stretching). The paradox was resolved when the functions of the molecular actors in contraction were finally understood. Here we review some of the classic experiments that led to this understanding
4. Actin-Myosin Interactions In Vitro: Dissections and Reconstitutions
An early experiment hinted at the interaction of actin and myosin in contraction. Homogenates of skeletal muscle were viscous. The viscous component was isolated and shown to contain a substance that was called actomyosin (acto, active; myosin, muscle substance). Under appropriate conditions, adding ATP to actomyosin preparations caused a decrease in viscosity. However, after the added ATP was hydrolyzed, the mixture became viscous again. Extraction of the nonviscous preparation (before it re-congealed and before the ATP was consumed) led to the biochemical separation of two the main substances we now recognize as the actin and myosin filaments of contraction. What’s more, adding these components back together reconstituted the viscous actomyosin extract (now referred to as actinomyosin to reflect its composition). And…, adding ATP to the reconstituted solution eliminated its viscosity. The ATP-dependent viscosity changes of actinomyosin solutions were consistent with an ATP-dependent separation of thick and thin filaments. Perhaps actin and myosin also separate in glycerinated muscles exposed to ATP, allowing them to stretch and relax.
The advent of electron microscopy provided further evidence of a role for ATP in both contraction and relaxation of skeletal muscle The purification of skeletal muscle actin (still attached to Z Lines) from myosin is cartooned below, showing what the separated components looked like in the electron microscope.

Next, when actin (still attached to Z-Lines) and myosin were mixed, electron microscopy of the resulting viscous material revealed thin filaments interdigitating with thick filaments. The result of this reconstitution experiment is shown below.
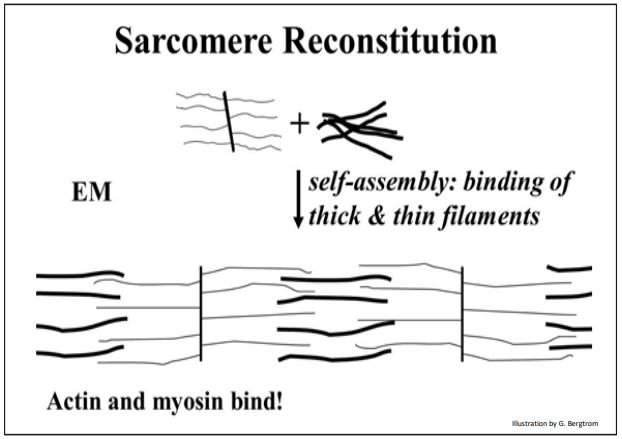
As expected, when ATP was added to these extracts, the solution viscosity dropped, and electron microscopy that the revealed thick (myosin) and thin (actin) filaments had again separated. The two components could again be isolated and separated by centrifugation.
In yet further experiments, actinomyosin preparations could be spread on over an aqueous surface, forming a film on the surface of the water. When ATP was added to the water, the film visibly “contracted”, pulling away from the edges of the vessel, reducing its surface area! Electron microscopy of the film revealed shortened sarcomere-like structures with closely spaced Z lines and short I bands…, further confirming the sliding filament model of muscle contraction.
332 In Vitro & Electron Microscope Evidence for a Sliding Filament Model
When actin and myosin were further purified from isolated actinomyosin, the thick myosin rods could be dissociated into large ~599Kd myosin monomers. Thus, thick filaments are massive polymers of myosin monomers! The molecular structure of myosin thick filaments is shown below.
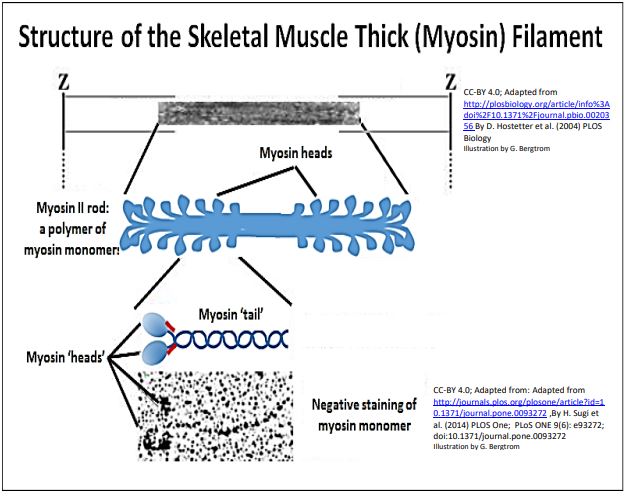
An early observation of isolated actin filaments was that they had no ATPase activity. On the other hand, while isolated myosin preparations did have an ATPase activity, they would only catalyze ATP hydrolysis very slowly compared to intact muscle fibers. Faster ATP hydrolysis occurred only if myosin filaments were mixed with microfilaments (either on, or detached from Z-lines). In the electron microscope, isolated myosin protein monomers appeared to have a double-head and single tail regions. Biochemical analysis showed that the myosin monomers themselves were composed of the two heavy chain and two pairs of light chain polypeptides shown in the illustration above
High magnification, high resolution electron micrographs and the corresponding illustration below show the component structures of myosin monomers.
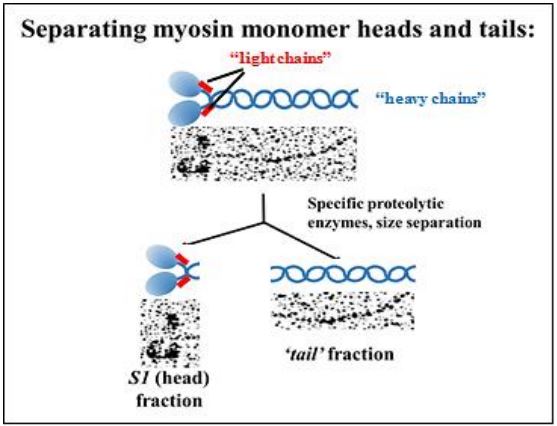
Proteolytic enzymes that hydrolyze peptide linkages only between specific amino acids, can ‘cut’ the heavy chains of myosin monomers into S1 (head) and tail fragments. Electron micrographs of these two fragments after separation by ultracentrifugation are shown above. S1 fragments were shown to have a slow ATPase activity, while the tails had none. The slow activity was not an artifact of isolation; mixing the S1 fraction with isolated actin filaments resulted in a higher rate of ATP hydrolysis. Clearly, myosin heads are ATPases that interact with actin microfilaments.
333 Thick Filament & Myosin Monomer Structure
The direct demonstration of an association of S1 myosin head fragments with rabbit smooth muscle actin microfilaments is shown below.

Just as for skeletal muscle, smooth muscle contraction is due to actin-myosin sliding, though smooth muscle is not striated and lacks sarcomere morphology; a white arrow in the micrograph points to one of several myosin (thick) filaments visible in the micrograph. The interaction of the S1 myosin heads with actin filaments dramatically alters their morphology. In this image, the diagonal stripes, or arrowhead-like appearance of the S1-actin binding all along actin filaments indicates that F-actin filaments are polar, with a plus (+) and a minus (–) end, as was expected.
The same “decoration” of microfilaments with arrowheads is seen when S1 heads (or even intact myosin monomers) bind to thin sections of skeletal muscle sarcomeres, preparations of actin still attached to the Z lines, and with isolated Factin preparations. These images are consistent with the requirement that myosin must bind to actin to achieve a maximum rate of ATPase activity during contraction.
The arrowheads on decorated actin still attached to Z lines always face in opposite directions, as shown below.
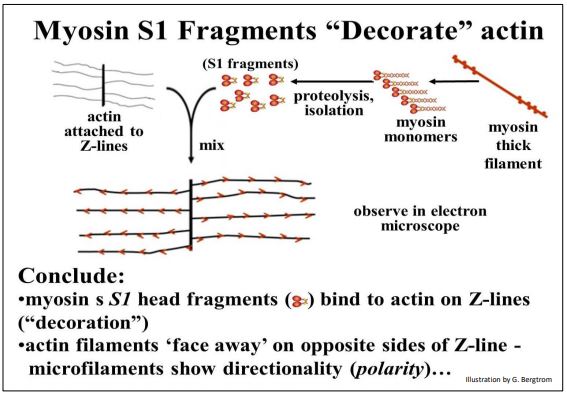
These opposing arrowheads, consistent with the sliding filament model of contraction in which bipolar thick filament pull actin filaments towards each other from opposite sides of the myosin rods, drawing the Z-lines closer together and shortening sarcomeres.
5. Allosteric Change and the Micro-Contraction Cycle
Whereas dynein and kinesin are motor proteins that ‘walk’ along microtubules, the myosin monomer is a motor protein that walks along microfilaments. In each case, these motor proteins are ATPases that use free energy of ATP hydrolysis to effect conformational changes that result in the walking, i.e., motility. In skeletal muscle, allosteric changes in myosin heads enable the myosin rods to do the walking along F-actin filaments
When placed in sequence such different myosin head conformations are likely the same as would occur during a micro-contraction cycle (illustrated below).
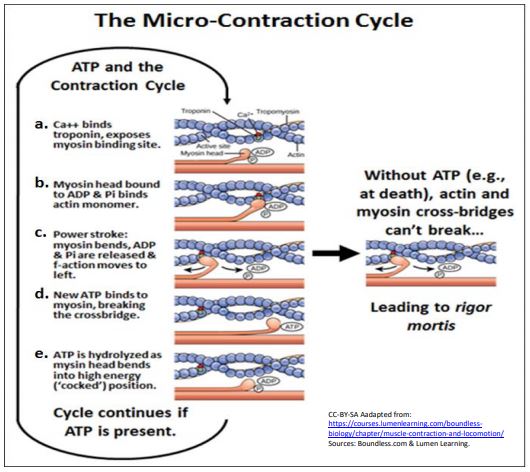
To help you follow the sequence, follow the small red dot on a single monomer in the actin filament. Here are the steps:
a. In the presence of Ca2+ ions, myosin binding sites on actin are open (Ca2+ - regulation of muscle contraction is discussed in more detail below).
b. Myosin heads with attached ADP and Pi bind to open sites on actin filaments.
c. The result of actin-myosin binding is an allosteric change in the myosin head, a bending of the hinge region, that pulls the attached microfilament (follow the red dot - it has moved from right to left!). This bit of micro-sliding of actin along myosin is the power stroke.
d. In its ‘bent’ conformation, the myosin head, still bound to an actin monomer in the F-actin, binds ATP, causing ADP and Pi to come off the myosin head and dissociating it from the actin.
e. Once dissociated from actin, myosin heads catalyze ATP hydrolysis, resulting in another conformational change. The head, still bound to ADP and Pi, has bent at its hinge, taking on a high-energy conformation that stores the energy of ATP hydrolysis.
f. The stored free energy is released during the power stroke. If Ca2+ has been removed, the myosins remain in the high-energy conformation of step e, until a release of Ca2+ again signals contraction.
Micro-contraction cycles of actin sliding along myosin continue as long as ATP is available. During repetitive micro-contraction cycles, myosin heads on the thick filaments pull actin filaments attached to Z-lines, bringing the Z lines closer to each other. The result is shortening of the sarcomere and ultimately, of muscle cells and the entire muscle. In the absence of ATP (as after the death of an organism), the micro-contraction cycle is interrupted. All myosin heads will remain bound to the actin filaments in the state of muscle contraction or relaxation (stretch) at the time of death. This is rigor mortis at the molecular level (see the illustration above). At the level of whole muscle, rigor mortis results in the inability to stretch or otherwise move body parts when ATP is, once and for all, depleted.
6. Resolving the Contraction Paradox
The myosin head micro-contraction cycle resolves the contraction paradox:
- ATP is necessary for muscle contraction: In step e in the illustration above, as ATP on myosin heads is hydrolyzed, the heads change from a low-energy to a high-energy conformation. The myosin heads can now bind to actin monomers (step b in the micro-contraction cycle). This results in of the power stroke (step c), where free energy released by an allosteric change in myosin, pulls the actin along the myosin, in effect causing a micro-shortening of the sarcomere, in other words, contraction!
- ATP is necessary for muscle relaxation: At the end of step c, myosin remains bound to actin until ATP can again bind to the myosin head. Binding of ATP in step d displaces ADP and inorganic phosphate (Pi) and breaks actinmyosin cross-bridges. A removal of Ca2+ from sarcomeres at the end of a contraction event blocks myosin binding sites on actin, while the rapid breakage of actin-myosin cross-bridges by ATP-myosin binding allows muscle relaxation and the sliding apart of the actin and myosin filaments (i.e., stretching). This leaves the myosin heads in the ‘cocked’ (high-energy) conformation, ready for the next round of contraction.
To summarize, ATP-myosin binding breaks actin-myosin cross-bridges. The muscle can then relax and stretch. Free energy of ATP hydrolysis, now stored in a high-energy myosin conformation, is released during the microcontraction power stroke
Electron microscopic examination of myosin monomer heads at different ionic strengths or when bound to antibodies (as shown below), provides visual evidence that myosin heads are flexible and can take on alternate stable conformations, as would be expected during the micro-contraction cycle.
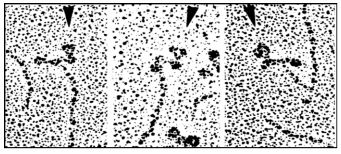
The arrow heads point to bound antibody molecules (immunoglobulins). For a video of conformational change in myosin monomers at Myosin heads in Action.
335 An Actin-Myosin Contraction Cycle Resolves the Contraction Paradox
336 Binding and Hydrolysis of ATP Changes Myosin Head Conformation
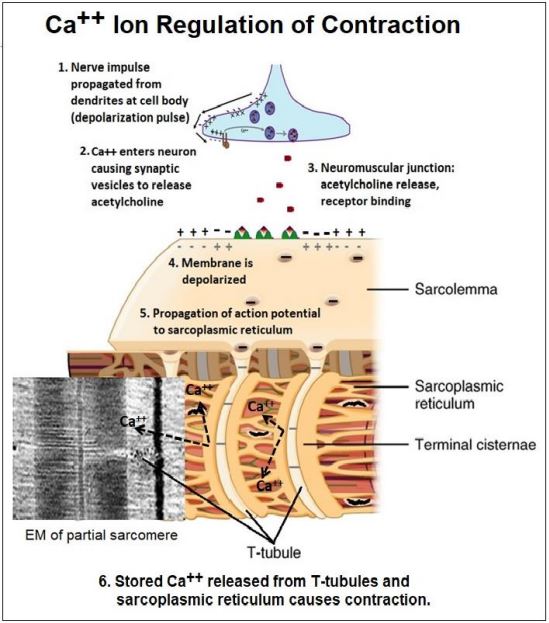
7. Ca2+ Ions Regulate Skeletal Muscle Contraction
Typically, the neurotransmitter acetylcholine released by a motor neuron binds to receptors on muscle cells to initiate contraction. In early experiments, Ca2+ was required, along with ATP, to get glycerinated skeletal muscle to contract. It was later demonstrated that Ca2+ ions were stored in the sarcoplasmic reticulum, the smooth endoplasmic reticulum of muscle cells. As we have seen, an action potential generated in the cell body of a neuron propagates along an axon to the nerve terminal, or synapse. In a similar fashion, an action potential generated at a neuromuscular junction travels along the sarcolemma (the muscle plasma membrane) to points where it is continuous with transverse tubules (T-tubules). The action potential then moves along the T-tubules and then along the membranes of the sarcoplasmic reticulum. This propagation of an action potential opens Ca2+ channels in the sarcoplasmic reticulum. The Ca2+ released bathes the sarcomeres of the myofibrils, allowing filaments to slide (i.e., contraction). The action potential at a neuromuscular junction that initiates contraction is summarized in the illustration below.
Ca2+ ions released from the sarcoplasmic reticulum bathe the myofibrils, where they bind to one of three troponin molecules to regulate skeletal muscle contraction. The three troponins and a tropomyosin molecule are bound to actin filaments.. Experiments using anti-troponin and anti-tropomyosin antibodies localize these proteins on thin filaments spaced at regular intervals in electron micrographs. The drawing below models this association of the troponin subunits and tropomyosin with the thin filaments.

In resting muscle, tropomyosin (a fibrous protein) lies along the actin filament where it covers up the myosin binding sites of seven G-actin subunits in the microfilament. In this conformation, troponin T (tropomyosin-binding troponin) and troponin I (inhibitory troponin) hold the tropomyosin in place. The ‘cross-section illustration below illustrates the role conformational changes in troponin C upon binding Ca++ in regulating contraction.
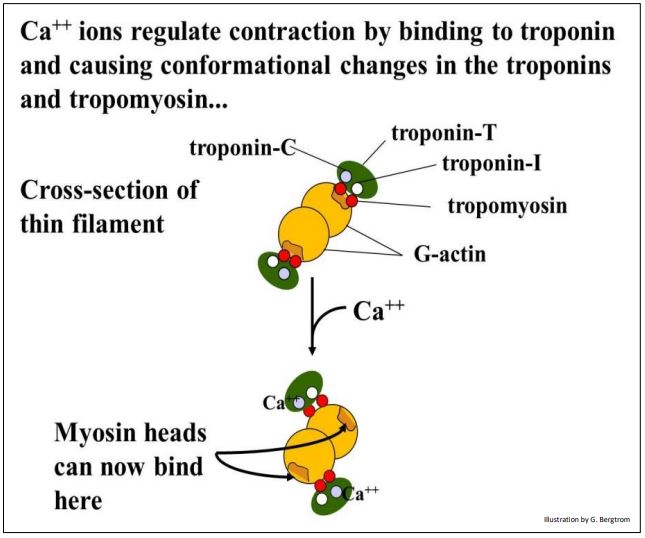
8. Muscle Contraction Generates Force
Contraction by ATP-powered sliding of thin along thick filaments generates force on the Z-lines. In three dimensions, the Z-lines are actually Z-disks) to which the actin thin filaments are attached. The protein \(\alpha \)-actinin in the Z-disks anchors the ends of the actin filaments to the disks so that when the filaments slide, the Z-disks are drawn closer, shortening the sarcomeres. Another Z-disk protein, desmin, is an intermediate filament organized around the periphery of Z-disks. Desmin connects multiple Z-disks in a myofibril. By keeping the Z-Disks in register, muscle cell, and ultimately, muscle contraction is coordinated. Finally, actin filaments at the ends of the cell must be connected to the cell membrane for a muscle cell to shorten during myofibril contraction. Several proteins, including syntrophins and dystrophin (another intermediate filament protein) anchor the free ends of microfilaments coming from Z-disks to the cell membrane. Still other proteins anchor the cell membrane in this region to the extracellular matrix (tendons) that are in turn, attached to bones! Force generated by myosin hydrolysis of ATP and the sliding of filaments in individual sarcomeres are thus transmitted to the ends of muscles to effect movement. If the name dystrophin sounds familiar, it should! The gene and its protein were named for a mutation that causes muscular dystrophy, resulting in a progressive muscle weakening.
338 Contraction Generates Force Against Z Disks and Cell Membranes
9. The Elastic Sarcomere: Do Myosin Rods Just Float in the Sarcomere?
In fact, myosin rods are anchored to proteins in the Z discs and M-lines. In 1954, R. Natori realized that when contracted muscle relaxes, it lengthens beyond its resting state, then shortening again to its resting length. He proposed that this elasticity must be due to a fiber in the sarcomere. Twenty-five years later, the elastic structure was identified as titin, a protein that holds several molecular records! The gene for titin contains the largest number of exons (363) of known proteins. After actin and myosin, titin is also the most abundant protein in muscle cells. At almost 4 x 106 Da, the aptly named titin is the also the largest known polypeptide. Extending from the Z discs to the M line of sarcomeres, titin coils around thick filaments along the way. Titin is anchored at Z-disks by \(\alpha \)-actinin and telethonin proteins. At the M-line, titin binds to myosin-binding protein C (MYBPC3) and calmodulin, among others (e.g., myomesin, obscurin and skelamin). Some if not all of these proteins must participate in keeping the myosin thick filaments positioned and in register in the sarcomere. This is similar to how Z-disks bind the ends of actin filaments to keep sarcomeres in register. The location of titin and several other sarcomere proteins is illustrated below.
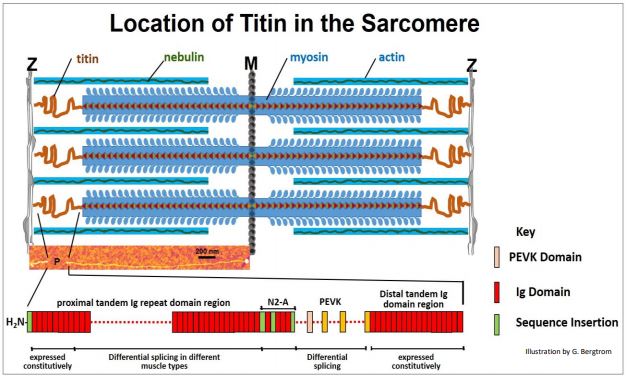
Coiled titin molecules (in red in the illustration) extend from the Z to M lines. The colorized electron micrograph of one extended titin molecule in the middle of the illustration above should convince you of the length (35,213 amino acids!) of this huge polypeptide! Titin’s elastic features are largely in the region labeled P in the micrograph, between Z discs and the myosin rods. The many domains of this P region are shown at the bottom of the illustration. With all the binding (and other) functions, you might expect that titin has many domains. It does! They include Ig (immunoglobulin) domains, fibronectin domains (not shown here), PEVK and N2A domains (that helps bind titin to \(\alpha \)-actinin in Z-disks). Which and how many Ig and/or PEVK domains are present in a particular muscle depends on which alternative splicing pathway is used to form a titin mRNA. Over a micron long, Titin functions as a molecular spring, as Natori predicted. Its coiled domains compress during contraction, passively storing some of the energy of contraction. When skeletal muscle relaxes, Ca2+ is withdrawn from the sarcomere, ATP displaces ADP from myosin heads and actin and myosin dissociate. The muscle then stretches, typically under the influence of gravity or an opposing set of muscles. However, during contraction, 244 individually folded protein domains of titin were compressed, and during relaxation, these domains de-compress; the stored energy of compression also helps to power relaxation. At the same time, titin connections limit the stretch so that a potentially overstretched muscle can ‘bounce’ back to its normal relaxed length.
In a particularly elegant experiment, R. Linke et al. provided a visual demonstration of myofiber elasticity consistent with the coiled spring model of titin structure. They made antibodies to peptide domains on either side of the PEVK domain of titin (N2A and I20-I22) and attached them to nanogold particles (which will appear as electron dense granules in transmission electron microscopy). Then individual myofibers were stretched to different lengths, fixed for electron microscopy and treated with the nanogold-linked antibodies. The antibodies localize to and define the boundaries of the titin PEVK domains. The image below does not show original immune-stained electron micrographs but show alternate sarcomere micrographs with simulated localization of nanogold particles, reflecting actual results.
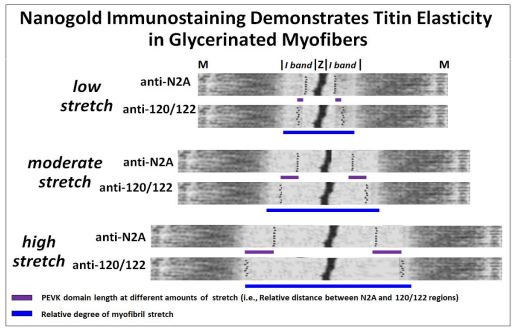
In the experiment, increased stretch lengthened the I bands on either side of Z lines of sarcomeres (blue bars). Likewise, the titin PEVK domains also lengthened as is evident from the increased distance between the nanogold-linked N2A and 120/122 antibodies that bind on either side PEVK domains. This demonstration of titin (and therefore sarcomere) elasticity) is consistent with the storage of some of the free energy of contraction when the molecule is compressed, and the passive release of that energy during relaxation. Since titin tethers thick filaments to Zdisks and M-lines, it also limits the amount of sarcomere stretch during relaxation. An animation from Linke’s lab is at http://www.titin.info/.
D. Non-muscle Microfilaments
Electron microscopy revealed that thin (~10 nm) filaments permeated the cytoskeleton of eukaryotic cells. These were suspected to be actin microfilaments. Microfilaments typically lie in the cortex of cells, just under the plasma membrane, where they support cell shape. These same microfilaments can also re-organize dynamically, allowing cells to change shape. A dramatic example of this occurs in dividing cells, during cytokinesis when the dividing cell forms a cleavage furrow in the middle of the cell. The cortical microfilaments slide past each other with the help of non-muscle myosin, progressively pinching the cell until it divides into two new cells.
To test whether these 10 nm ‘microfilaments’ were in fact actin, intact myosin monomers or S1 myosin head fragments were placed atop electron micrographs of many different cell types. When viewed in the electron microscope, such preparations always revealed that the 10 nm microfilaments were decorated with arrowheads, just like S1 fragment decorated muscle cell actin or Z line-bound actin! Clearly, these cytoplasmic microfilaments are a form of F-actin. In the example shown below, cells in cytokinesis were treated with S1 myosin head fragments.
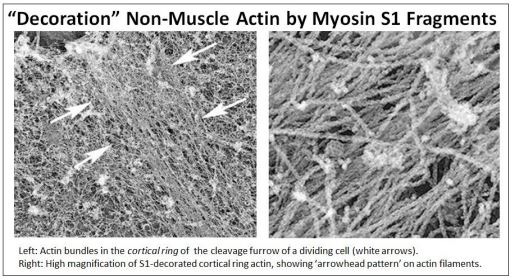
See the role of cortical filaments in cytokinesis at Cortical Actin Filament Action in Cytokinesis. Of course, actin microfilaments are involved in all manner of cell motility in addition to their role in cell division. They enable cell movement and cytoplasmic streaming inside cells. And while they give intestinal microvilli strength, they even enable microvilli to move independent of the passive pressures of peristalsis.
Other examples of microfilaments in cell motility include the ability of amoeba and other phagocytic cells to extend pseudopodia to engulf food or foreign particles (e.g., bacteria), respectively. Similarly, when fibroblast cells move along surfaces, they extend thin filipodia into the direction of movement by assembling actin bundles along the axis of cell movement. Actin stress fibers that help to maintain cell shape fluorescence green in the immunofluorescence micrograph below (left panel). The dual roles of actin in fibroblast movement are also illustrated (below right).
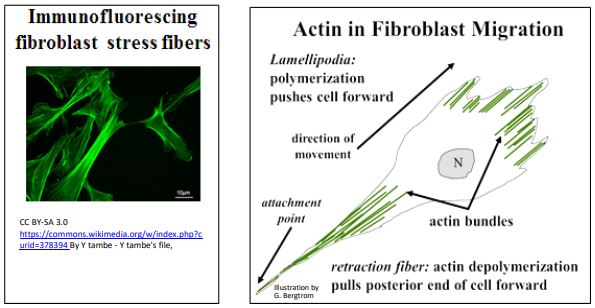
As we saw for microtubule-mediated cell motility, some actin-mediated motility may be primarily based on actin assembly and disassembly, as in the extension of filipodia at the moving front of a fibroblast. As the fibroblast moves forward, a retraction fiber at the hind-end of the cell remains attached to the surface (substratum) along which it is migrating. Eventually however, actin-myosin interactions (in fact, sliding) causes retraction of most of this ‘fiber’ back into the body of the cell.
Movements mediated by stress fibers may also explain the cytoplasmic streaming that distributes cellular components and nutrients throughout a cell. The movements of both involve actin-myosin interactions. Studies of non-muscle cell motility suggest the structure and interacting molecular components of stress fibers. They reveal overlapping myosin and actin filaments that slide during movement, as illustrated below.
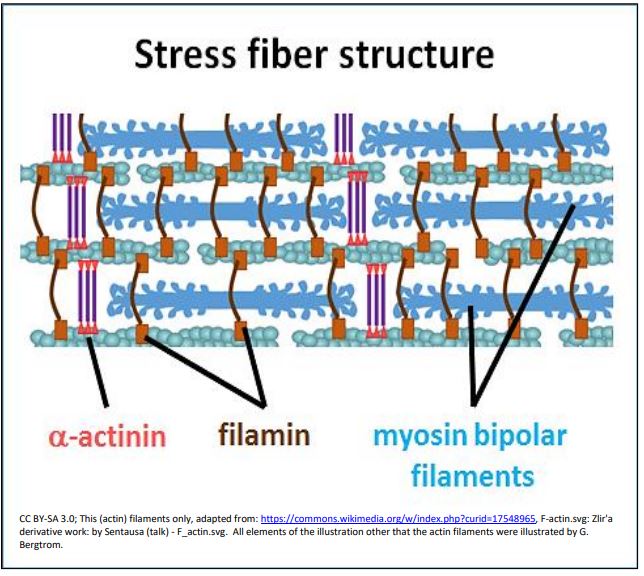
Filamin in this drawing is shown holding actin filaments together at an angle, while \(\alpha \)- actinin also helps to bundle the actin (thin) filaments. Titin (not shown) also seems to be associated with stress fibers. However, unlike highly organized skeletal muscle sarcomeres, the proteins and filaments in stress fibers are not part of Z- or M-line superstructures. Could such less-organized non-muscle stress fiber filament bundles be the evolutionary predecessor to sarcomeres in muscle cells?
E. Actins and Myosins are Encoded by Large Gene Families
Actins may be the most abundant protein in cells! At least six different actin isoforms encoded by a large actin gene family have nearly identical amino acid sequences, all of which are involved in cytoskeletal function. The \(\beta \)-actin isoform predominates. Genes for some isoforms are expressed in a cell-specific manner.
Are all actin isoforms functionally significant? Myosin monomers (or S1 heads) decorate virtually all actins. This makes one wonder if any one actin is an adaptation, however subtle, such that the absence of one isoform would pose a significant threat to the survival of an organism? Since amino acid sequence differences between actins would not predict dramatically different protein function, could they underlie some as yet unknown physiological advantage to different cells? In mice, the loss of a \(\gamma \)-actin gene has little effect on the organism, while loss of the \(\beta \)-actin gene in mice is lethal at embryonic stages. In contrast, studies show that a mutated \(\beta \)-actin gene in humans correlate with delayed development and later neurological problems (e.g., epilepsy), kidney and heart abnormalities, but is not lethal. In fact, people with such mutations can lead nearly normal, healthy lives (Beta-Actin Gene Mutations and Disease).
Like the actins, myosin genes encoding variant isoforms comprise a large eukaryotic gene family. All isoforms have ATPase activity and some are clearly involved in cell motility. Unique functions are not yet known for other isoforms, but different myosin monomers can decorate actin, and myosins from one species can decorate actin filaments of other species, even across wide phylogenetic distances.
F. Intermediate Filaments - an Overview
These 10 nm filaments are proteins with extended secondary structure that in fact, do not readily fold into tertiary structures, and they have no enzymatic activity. Recall their intercellular location in desmosomes where they firmly bind cells together to confer tensile strength to tissues. Within cells, intermediate filaments permeate the cells where they participate in regulating and maintaining cell shape. Recall their role in anchoring actin to either Z-disks or plasma membrane plaques in muscle cells, transmitting the forces of contraction to the shortening of the sarcomeres and then to the actual shortening of a muscle. The extracellular keratins that make up fur, hair, fingernails and toenails, are proteins related to intermediate filaments. Unlike intracellular intermediate filaments, keratins are bundles of rigid, insoluble extracellular proteins that combine to align to form stable, unchanging secondary structures. Finally, lamins are intermediate filaments that make up structural elements of the nuclear lamina, a kind of nucleoskeleton.
CMB3e 492 As we saw earlier, intermediate filament subunits have a common structure consisting of a pair of monomers, each with globular domains at their C- and N-terminal ends, separated by coiled rod regions. Monomers are non-polar; i.e., unlike microtubules and actin filaments, they do not have ‘plus’ and ‘minus’ ends. The basic unit of intermediate filament structure is a dimer of monomers. Dimers further aggregate to form tetramers and larger filament bundles. Like microtubules and actin filaments, intermediate filament bundles can disassemble and reassemble as needed when cells change shape. Unlike microtubules and actin, intermediate filaments can stretch, a property conferred by the coiled rod regions of the filaments. This should be reminiscent of titin molecules! The structural features and elasticity of intermediate filaments is illustrated in the cartoon below.
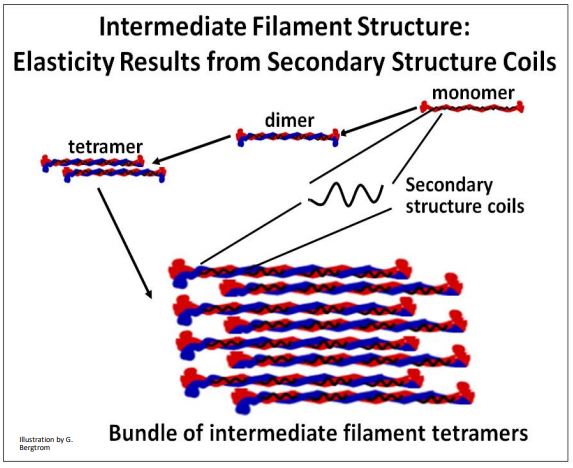
In the bundled intermediate filaments that permeate the cytoplasm of cells, the ability to stretch contributes to the viscosity of cytoplasm, and is even called viscoelasticity. This elastic property is thought to allow actins and microtubules a degree of freedom of movement of cells, and within the cytoplasm of cells.