
15.1: Overview
- Page ID
- 16506

We start this chapter by looking at technologies that led to genetic engineering. The ability of make recombinant DNA is such a seminal technology that just realizing it could be done and then doing it in a test tube for the first time earned Paul berg a half-share in the 1980 Nobel Prize in Chemistry (the other half was shared by Walter Gilbert and Frederick Sanger for studies that enabled efficient DNA sequencing). First we’ll look at cDNA synthesis, the synthesis of DNA copies from RNA, something retroviruses routinely do as part of the pathway of their reproduction. The retrovirus injects its RNA into target cells where it transcribes a reverse transcriptase enzyme. The enzyme reversetranscribes a copy DNA (the cDNA) complementary to the viral RNA. The First steps in retroviral infection is summarized in the illustration below.
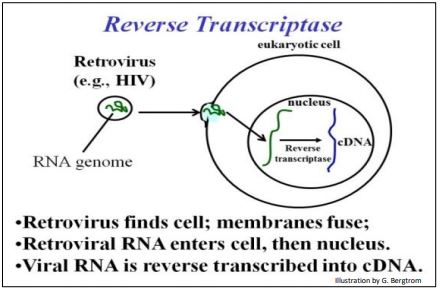
The same reverse transcriptase enzyme makes a double stranded cDNA, or (ds)cDNA, which then replicates. These cDNAs are transcribed into new viral RNA genomes and mRNAs for viral proteins. The latter encapsulate the RNA genomes into new viruses. Reverse transcriptase is now a laboratory tool, used to reverse transcribe cDNA from virtually any RNA sequence. It, along with many viral, bacterial and even eukaryotic enzymes and biomolecules, are now part of our recombinant DNA and genetic engineering toolkit.
We will see how a cDNA library is made and screened for a cDNA clone, and how a cloned cDNA can fish an entire gene out of a genomic library. Next we will see how the polymerase chain reaction (PCR) can produce (amplify) millions of copies of a single gene (or other DNA sequence) from as little DNA as is found in a single cell. Apart from its well-publicized use in forensics, PCR is another important laboratory tool for fetching, amplifying and studying sequences of interest. These venerable technologies illustrate important principles of cloning and sequence analysis. Of course, the analysis of traditionally cloned and amplified DNA sequences has been used to study the evolution and expression of individual genes. And sometimes we are misled! For example, knowing that a genetic mutation is associated with an illness usually leads to a search for how the mutation might cause the illness. But, as researchers in any discipline keep warning us, correlation is not causation! In fact, we know that many phenotypes, including genetic disease, are not the result of a single mutant gene. Autism is just one example.
The newer fields of genomics and proteomics leverage a growing battery of powerful tools to study many genes and their regulatory networks at the same time. The molecular networking made possible by genomics and proteomics (and other colorful holistic terms we’ll discuss later) promise to get us past naïve and often incorrect notions of causation. We may be soon able to identify many correlations that might sum up to causation or propensity to genetic illness.